

Identification of constrained sequence elements across 239 primate genomes
- PMID: 38030727
- PMCID: PMC10808062
- DOI: 10.1038/s41586-023-06798-8
Identification of constrained sequence elements across 239 primate genomes
Abstract
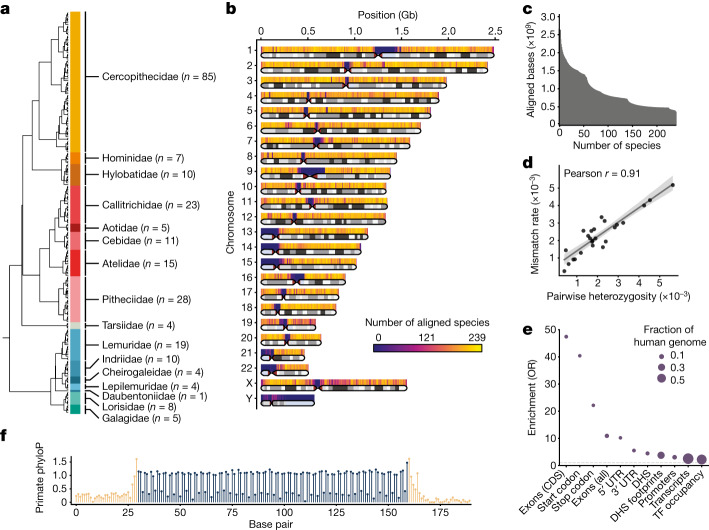
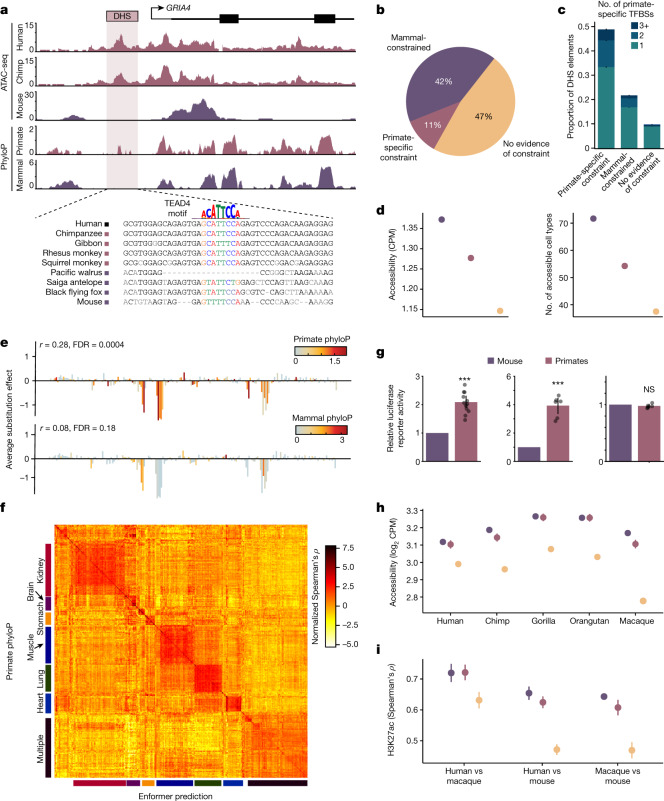
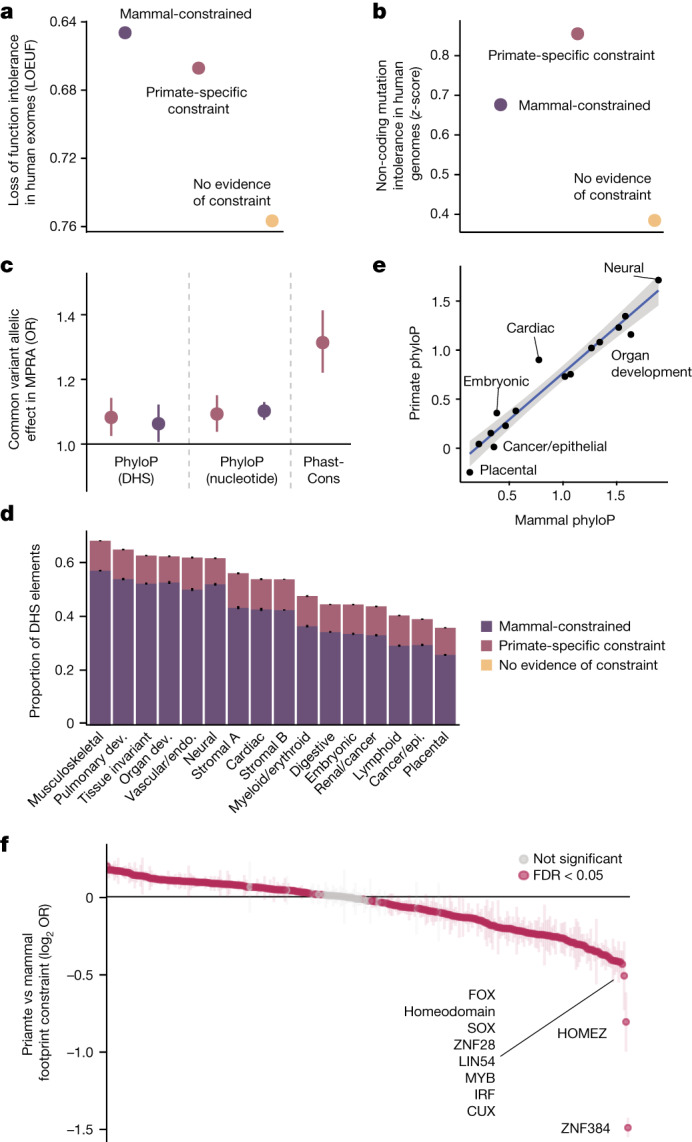
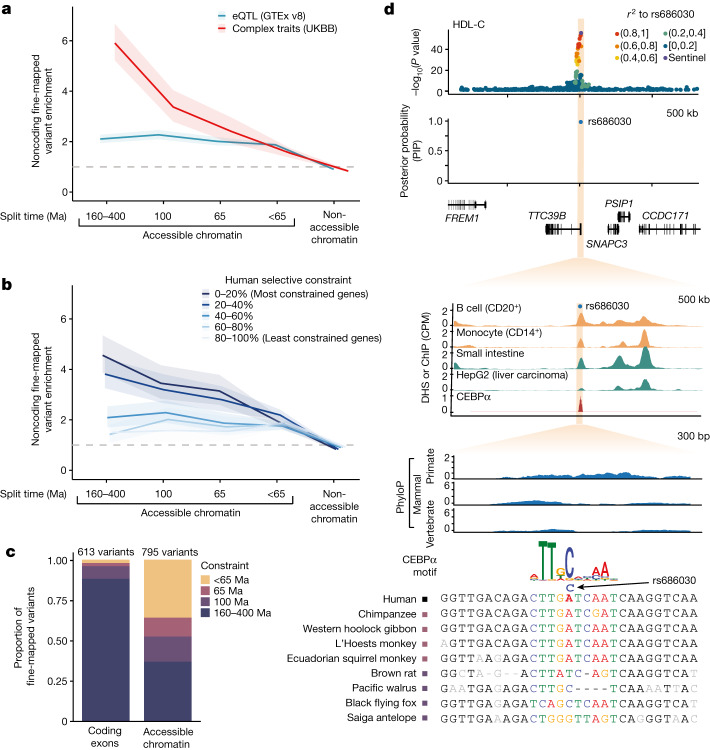
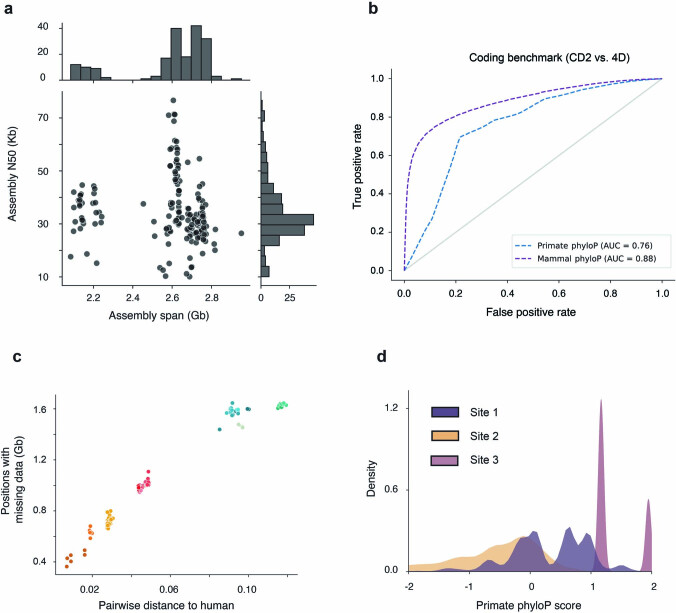
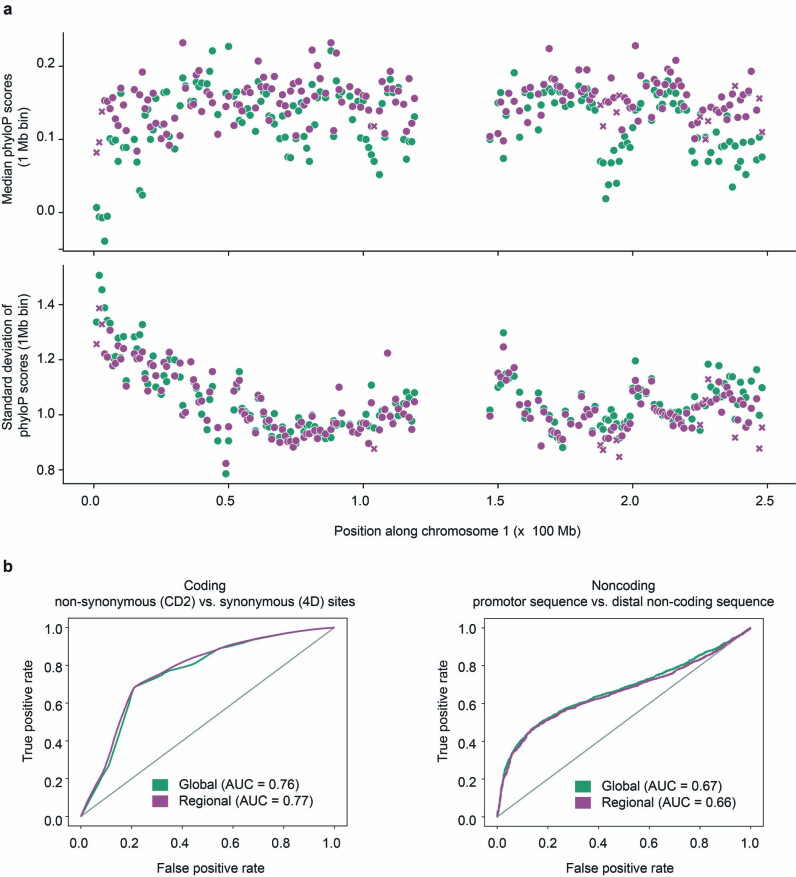
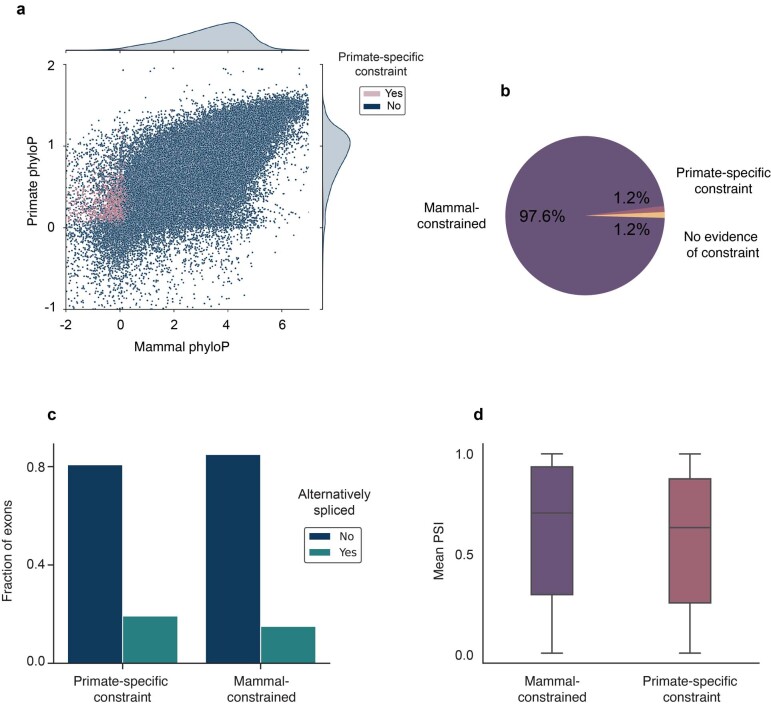
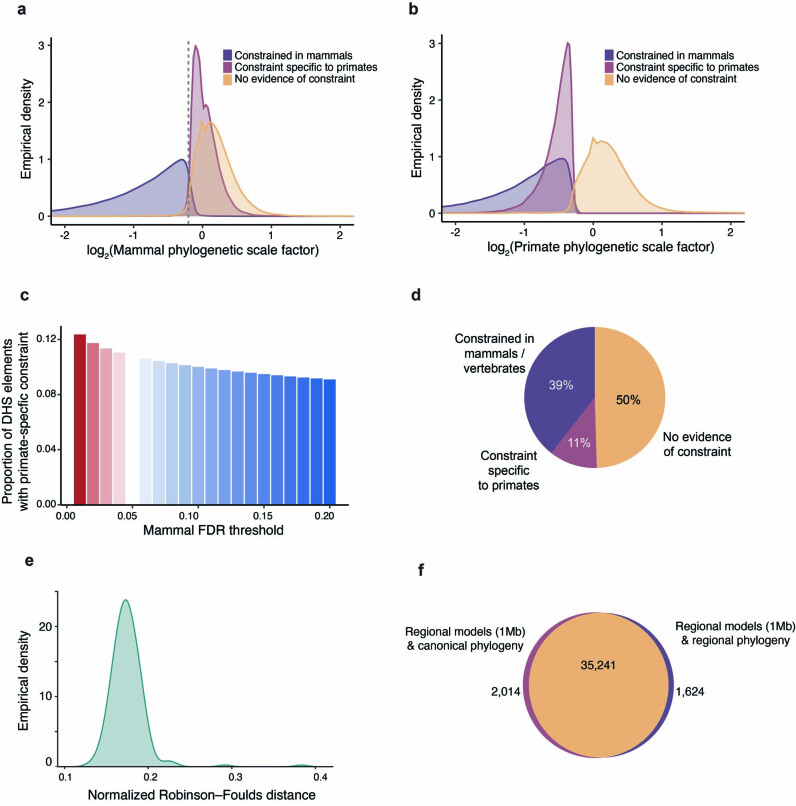
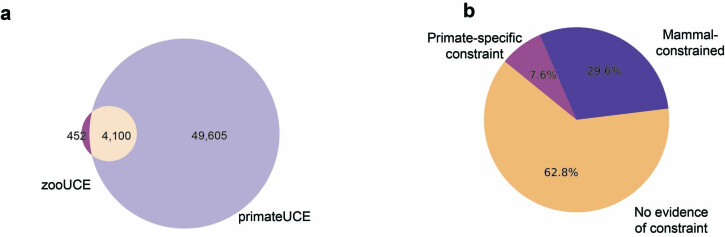
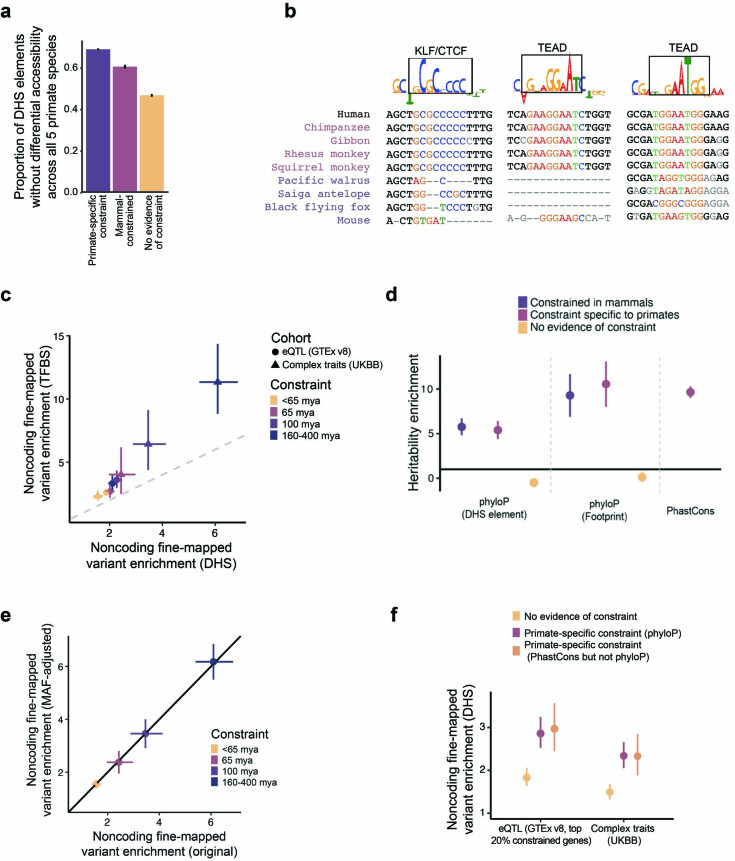
Noncoding DNA is central to our understanding of human gene regulation and complex diseases1,2, and measuring the evolutionary sequence constraint can establish the functional relevance of putative regulatory elements in the human genome3-9. Identifying the genomic elements that have become constrained specifically in primates has been hampered by the faster evolution of noncoding DNA compared to protein-coding DNA10, the relatively short timescales separating primate species11, and the previously limited availability of whole-genome sequences12. Here we construct a whole-genome alignment of 239 species, representing nearly half of all extant species in the primate order. Using this resource, we identified human regulatory elements that are under selective constraint across primates and other mammals at a 5% false discovery rate. We detected 111,318 DNase I hypersensitivity sites and 267,410 transcription factor binding sites that are constrained specifically in primates but not across other placental mammals and validate their cis-regulatory effects on gene expression. These regulatory elements are enriched for human genetic variants that affect gene expression and complex traits and diseases. Our results highlight the important role of recent evolution in regulatory sequence elements differentiating primates, including humans, from other placental mammals.
© 2023. The Author(s).
Conflict of interest statement
L.F.K.K., J.C.U., S.R., M.A., L.S., A.J.C., H.G., A.K., F.A., J.G.S. and K.K.-H.F. were employees of Illumina Inc. as of the initial submission of this manuscript.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
