

Adipose Triglyceride Lipase Is a Therapeutic Target in Advanced Prostate Cancer That Promotes Metabolic Plasticity
- PMID: 38038968
- PMCID: PMC10939928
- DOI: 10.1158/0008-5472.CAN-23-0555
Adipose Triglyceride Lipase Is a Therapeutic Target in Advanced Prostate Cancer That Promotes Metabolic Plasticity
Abstract
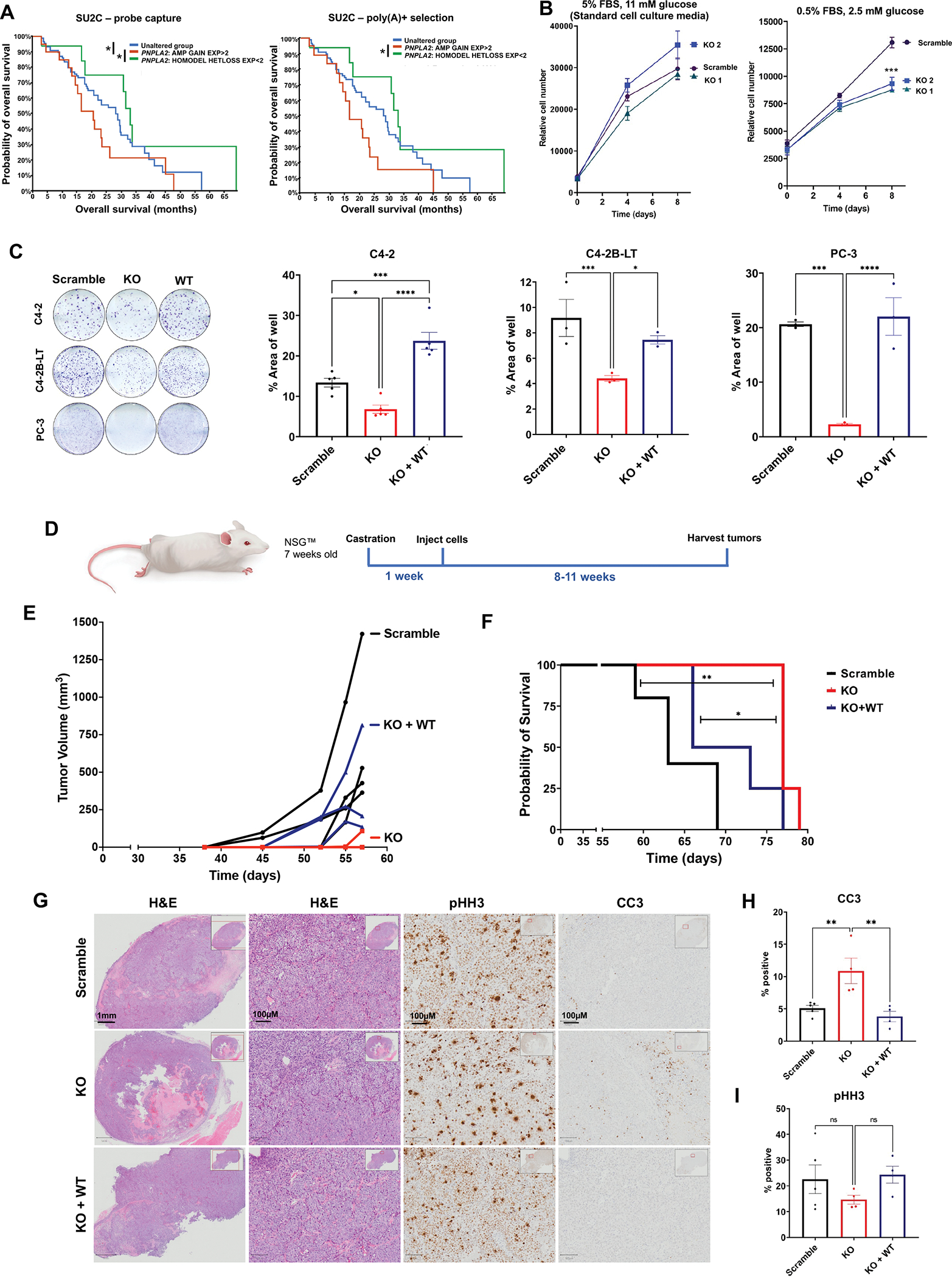
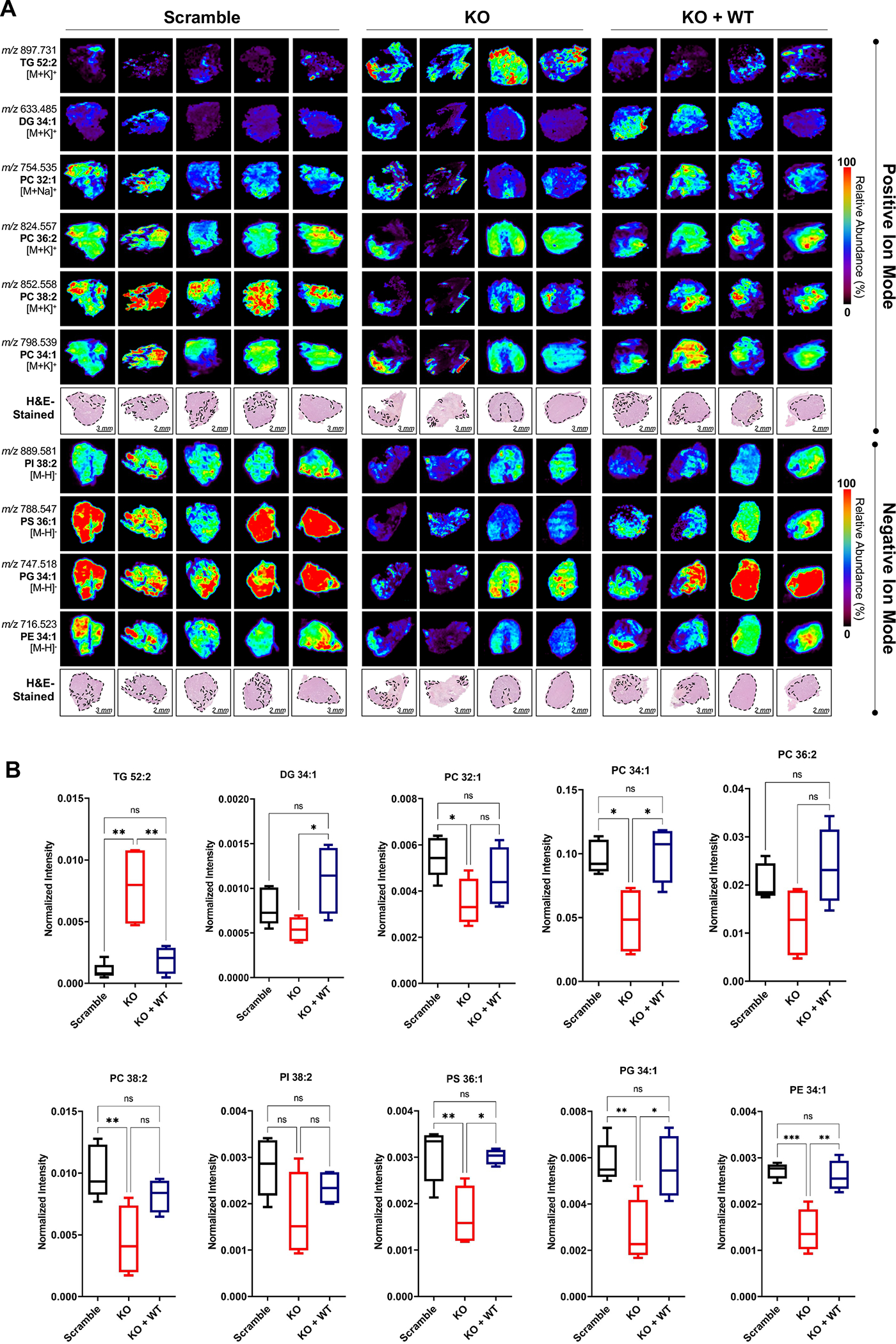
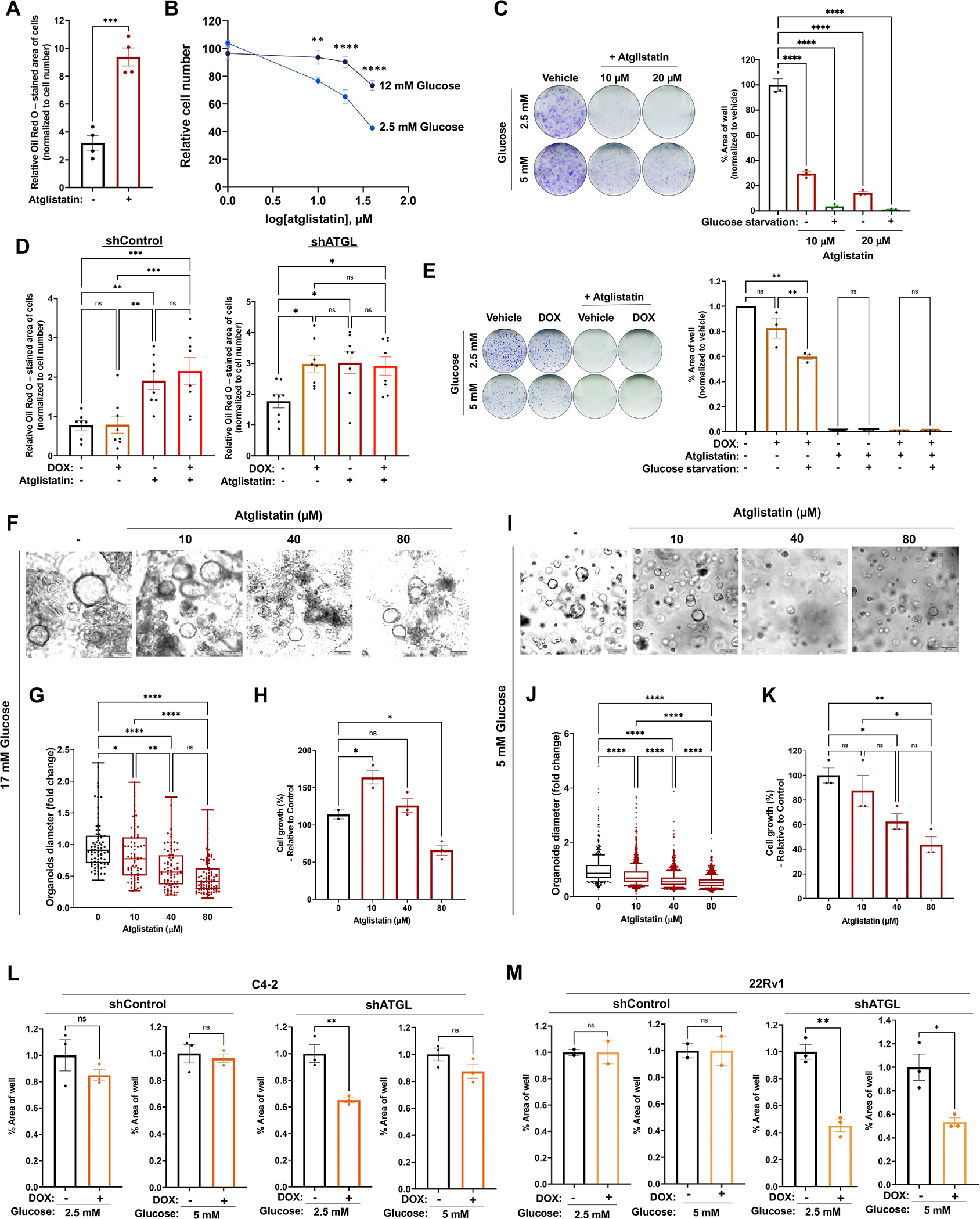
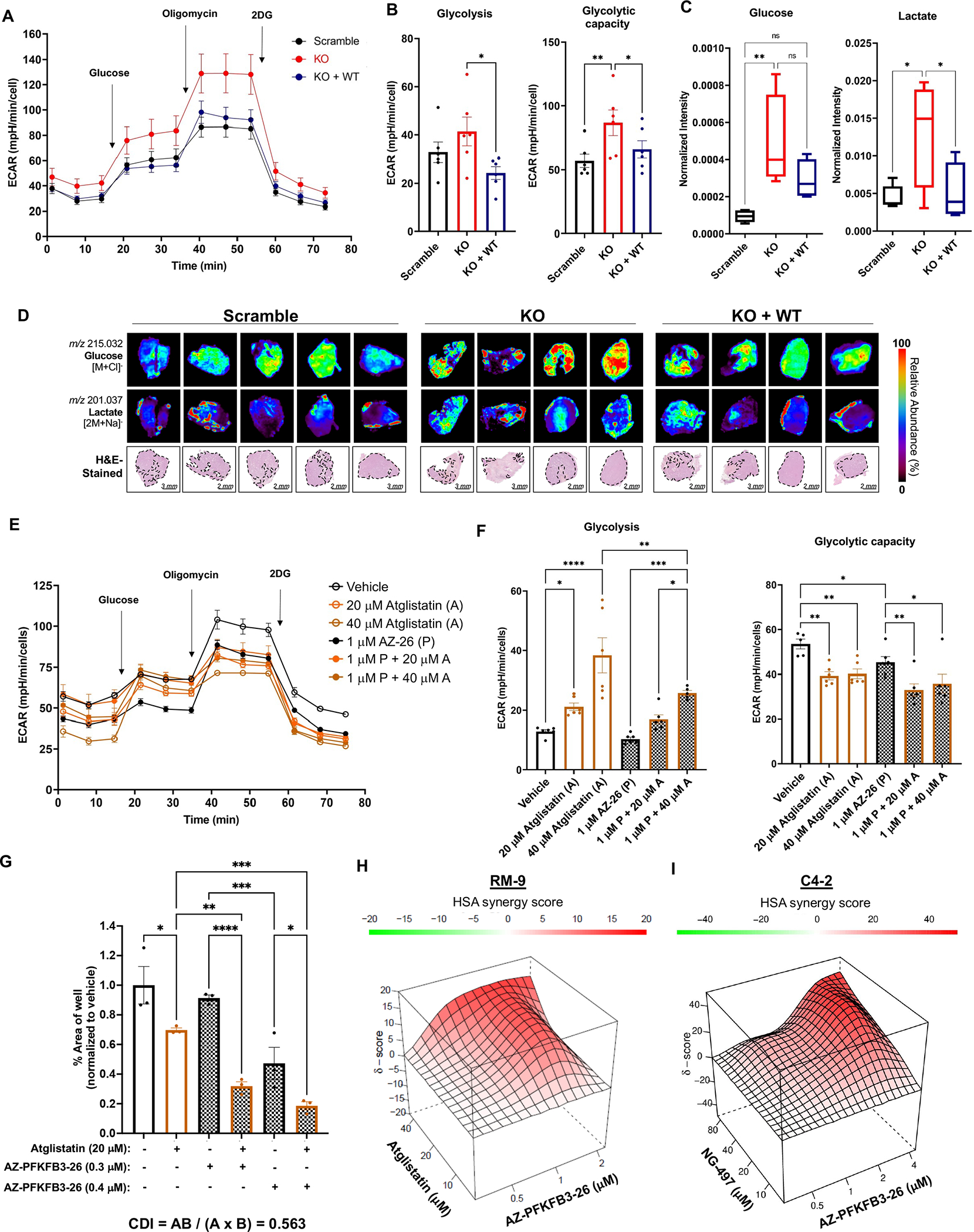
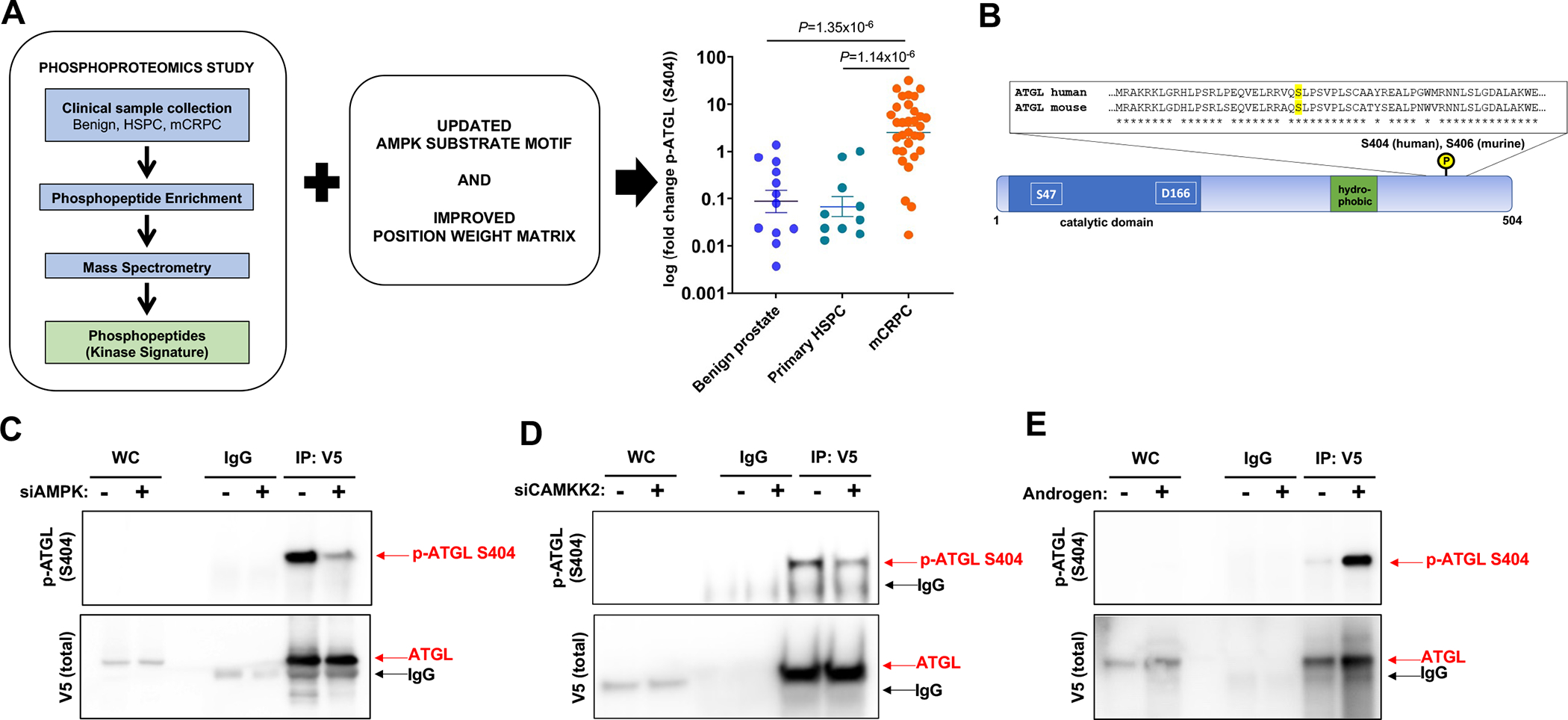
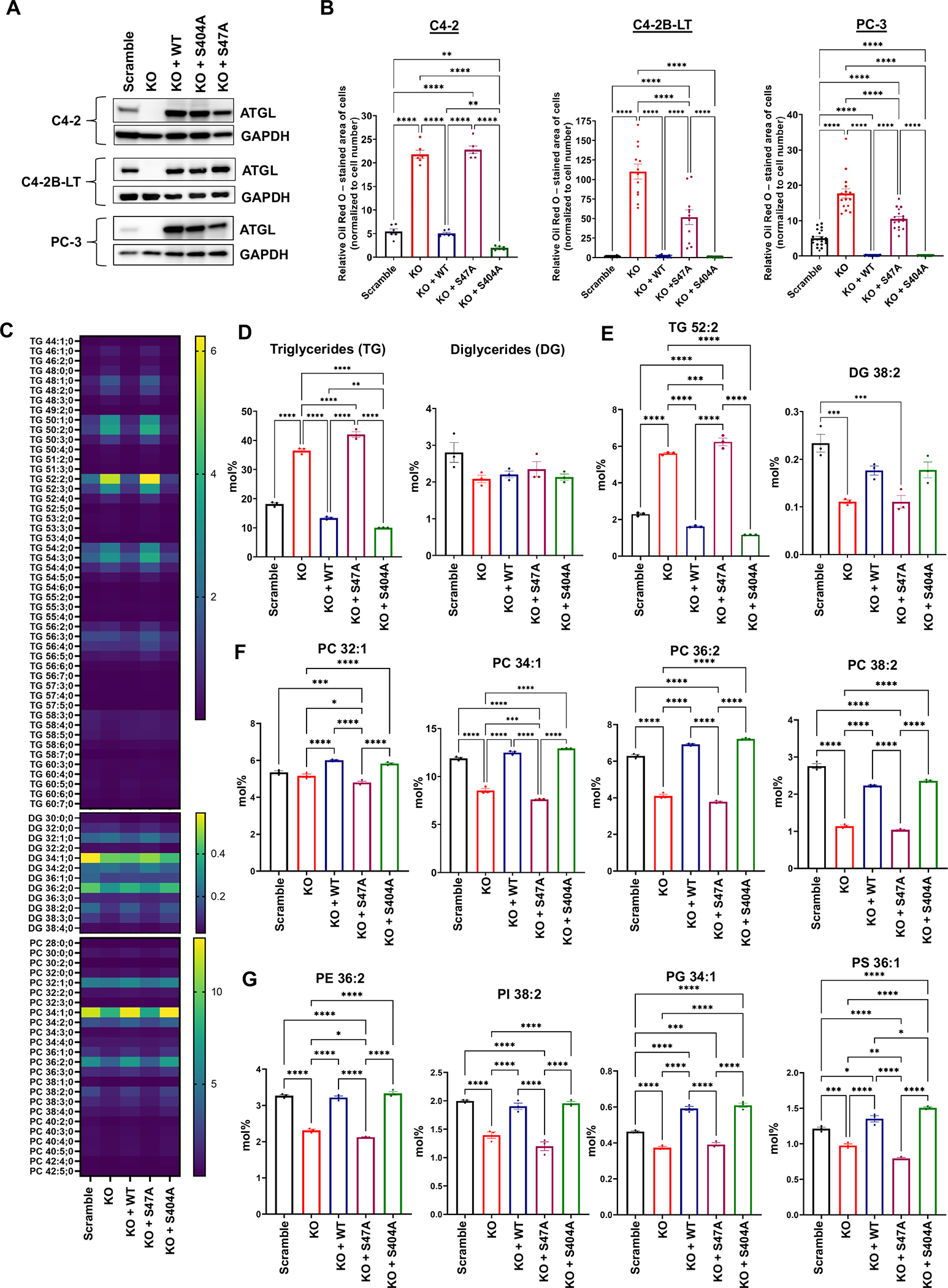
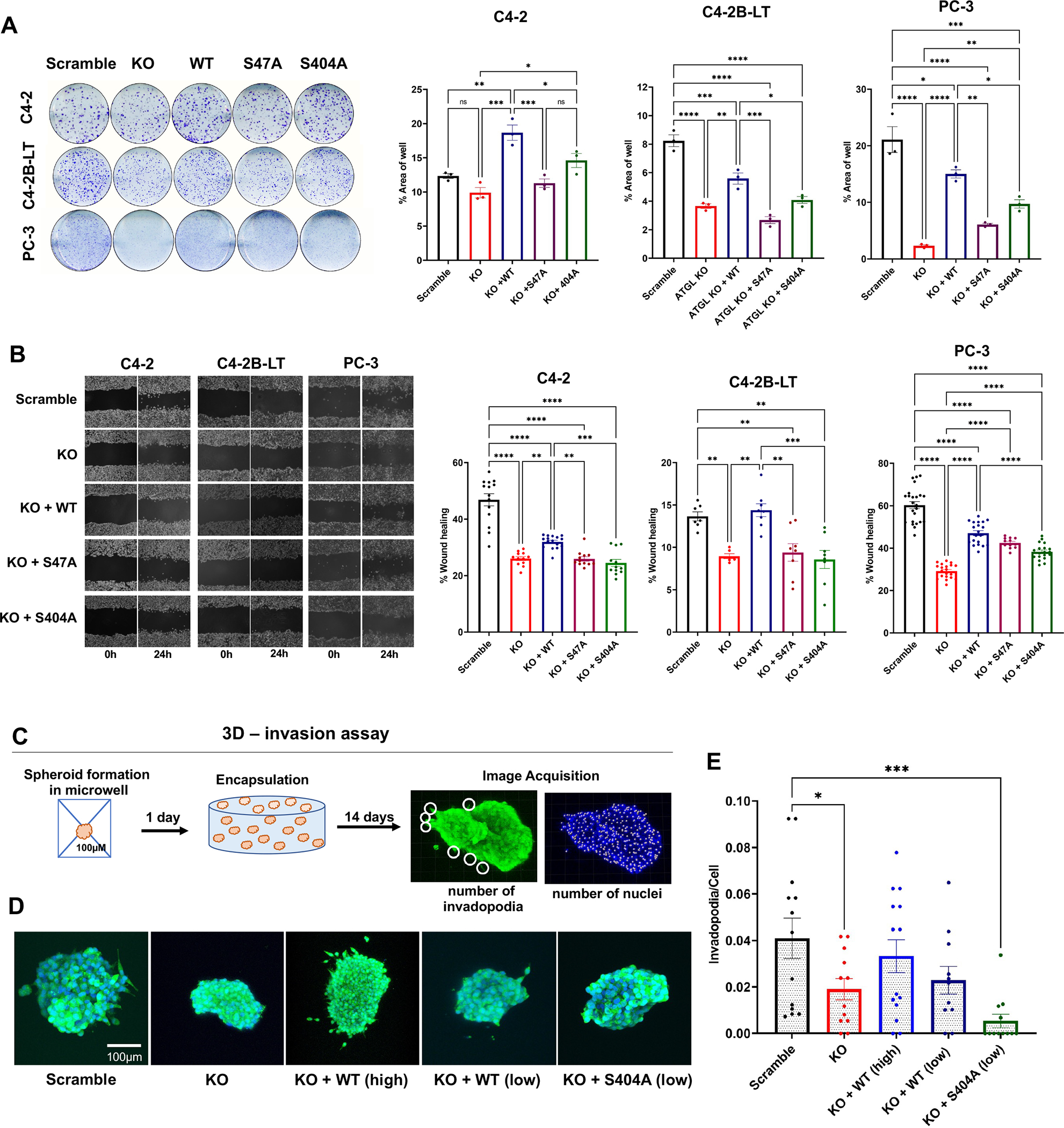
Lipid metabolism plays a central role in prostate cancer. To date, the major focus has centered on de novo lipogenesis and lipid uptake in prostate cancer, but inhibitors of these processes have not benefited patients. A better understanding of how cancer cells access lipids once they are created or taken up and stored could uncover more effective strategies to perturb lipid metabolism and treat patients. Here, we identified that expression of adipose triglyceride lipase (ATGL), an enzyme that controls lipid droplet homeostasis and a previously suspected tumor suppressor, correlates with worse overall survival in men with advanced, castration-resistant prostate cancer (CRPC). Molecular, genetic, or pharmacologic inhibition of ATGL impaired human and murine prostate cancer growth in vivo and in cell culture or organoids under conditions mimicking the tumor microenvironment. Mass spectrometry imaging demonstrated that ATGL profoundly regulates lipid metabolism in vivo, remodeling membrane composition. ATGL inhibition induced metabolic plasticity, causing a glycolytic shift that could be exploited therapeutically by cotargeting both metabolic pathways. Patient-derived phosphoproteomics identified ATGL serine 404 as a target of CAMKK2-AMPK signaling in CRPC cells. Mutation of serine 404 did not alter the lipolytic activity of ATGL but did decrease CRPC growth, migration, and invasion, indicating that noncanonical ATGL activity also contributes to disease progression. Unbiased immunoprecipitation/mass spectrometry suggested that mutation of serine 404 not only disrupts existing ATGL protein interactions but also leads to new protein-protein interactions. Together, these data nominate ATGL as a therapeutic target for CRPC and provide insights for future drug development and combination therapies.
Significance: ATGL promotes prostate cancer metabolic plasticity and progression through both lipase-dependent and lipase-independent activity, informing strategies to target ATGL and lipid metabolism for cancer treatment.
©2023 American Association for Cancer Research.
Conflict of interest statement
Figures
References
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell 2011;144:646–74 - PubMed
-
- Watt MJ, Clark AK, Selth LA, Haynes VR, Lister N, Rebello R, et al. Suppressing fatty acid uptake has therapeutic effects in preclinical models of prostate cancer. Sci Transl Med 2019;11 - PubMed
-
- Zimmermann R, Strauss JG, Haemmerle G, Schoiswohl G, Birner-Gruenberger R, Riederer M, et al. Fat mobilization in adipose tissue is promoted by adipose triglyceride lipase. Science 2004;306:1383–6 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
