

Design and Engineering of Light-Induced Base Editors Facilitating Genome Editing with Enhanced Fidelity
- PMID: 38039441
- PMCID: PMC10837352
- DOI: 10.1002/advs.202305311
Design and Engineering of Light-Induced Base Editors Facilitating Genome Editing with Enhanced Fidelity
Abstract
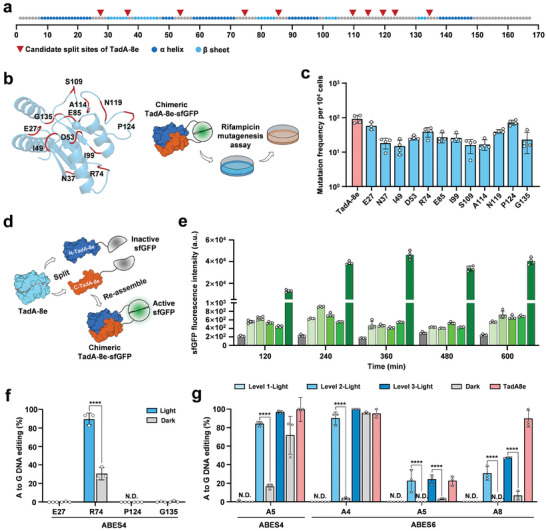
Base editors, which enable targeted locus nucleotide conversion in genomic DNA without double-stranded breaks, have been engineered as powerful tools for biotechnological and clinical applications. However, the application of base editors is limited by their off-target effects. Continuously expressed deaminases used for gene editing may lead to unwanted base alterations at unpredictable genomic locations. In the present study, blue-light-activated base editors (BLBEs) are engineered based on the distinct photoswitches magnets that can switch from a monomer to dimerization state in response to blue light. By fusing the N- and C-termini of split DNA deaminases with photoswitches Magnets, efficient A-to-G and C-to-T base editing is achieved in response to blue light in prokaryotic and eukaryotic cells. Furthermore, the results showed that BLBEs can realize precise blue light-induced gene editing across broad genomic loci with low off-target activity at the DNA- and RNA-level. Collectively, these findings suggest that the optogenetic utilization of base editing and optical base editors may provide powerful tools to promote the development of optogenetic genome engineering.
Keywords: base editor; low off-target effect; optogenetics; spatiotemporal control.
© 2023 The Authors. Advanced Science published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources