

Eyes open on stem cells
- PMID: 38039972
- PMCID: PMC10724227
- DOI: 10.1016/j.stemcr.2023.10.023
Eyes open on stem cells
Abstract
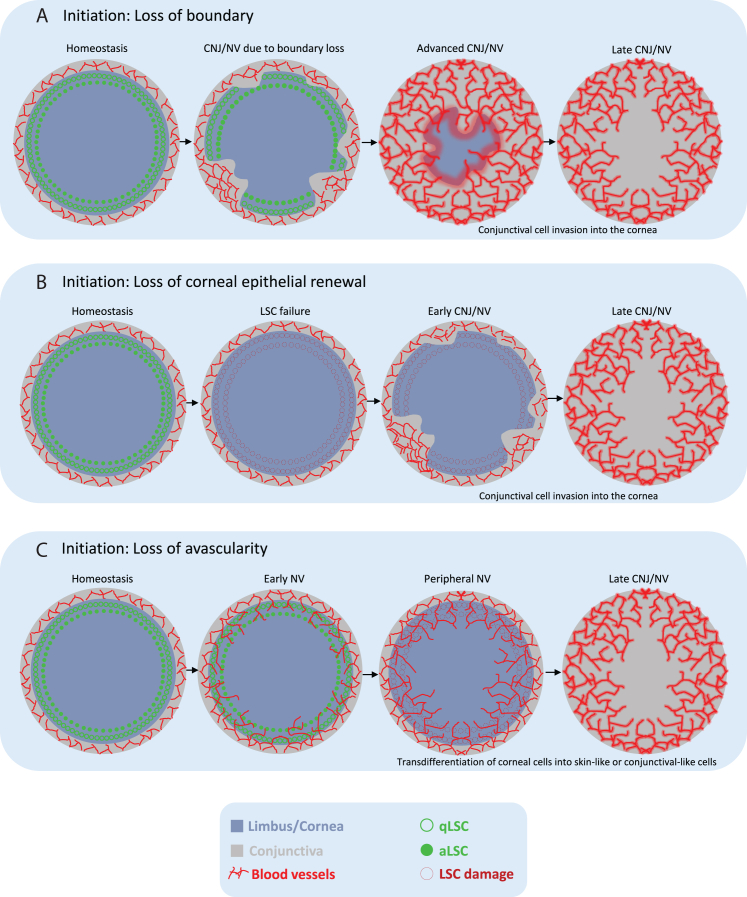
Recently, the murine cornea has reemerged as a robust stem cell (SC) model, allowing individual SC tracing in living animals. The cornea has pioneered seminal discoveries in SC biology and regenerative medicine, from the first corneal transplantation in 1905 to the identification of limbal SCs and their transplantation to successfully restore vision in the early 1990s. Recent experiments have exposed unexpected properties attributed to SCs and progenitors and revealed flexibility in the differentiation program and a key role for the SC niche. Here, we discuss the limbal SC model and its broader relevance to other tissues, disease, and therapy.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






References
-
- Altshuler A., Amitai-Lange A., Tarazi N., Dey S., Strinkovsky L., Hadad-Porat S., Bhattacharya S., Nasser W., Imeri J., Ben-David G., et al. Discrete limbal epithelial stem cell populations mediate corneal homeostasis and wound healing. Cell Stem Cell. 2021;28:1248–1261.e8. doi: 10.1016/j.stem.2021.04.003. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
