

A two-kinesin mechanism controls neurogenesis in the developing brain
- PMID: 38040957
- PMCID: PMC10692124
- DOI: 10.1038/s42003-023-05604-5
A two-kinesin mechanism controls neurogenesis in the developing brain
Abstract
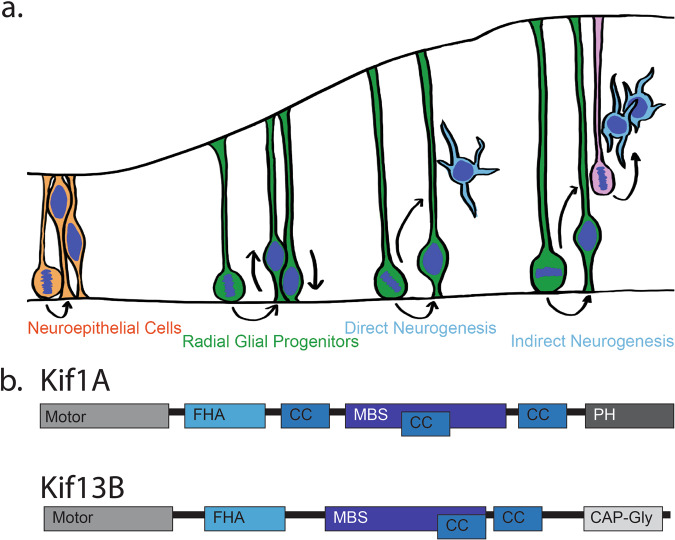
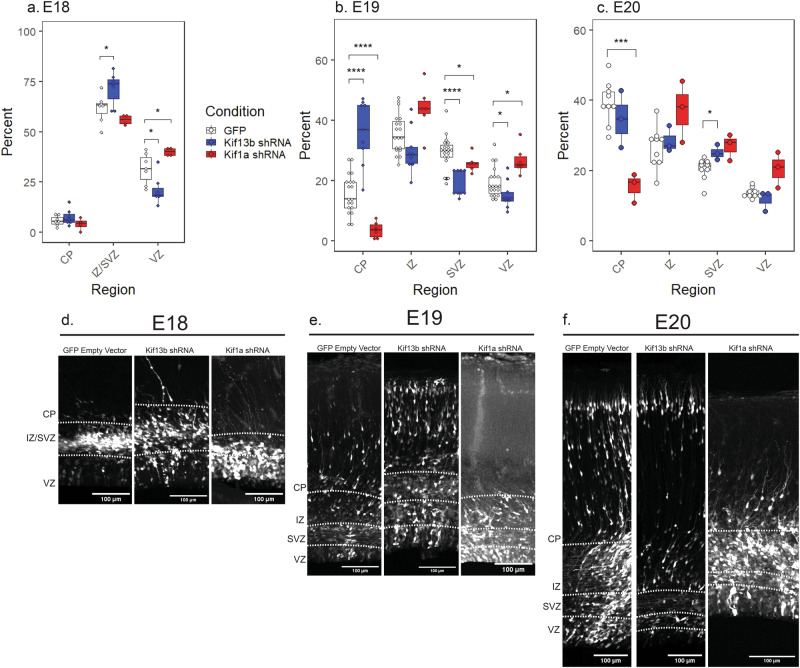
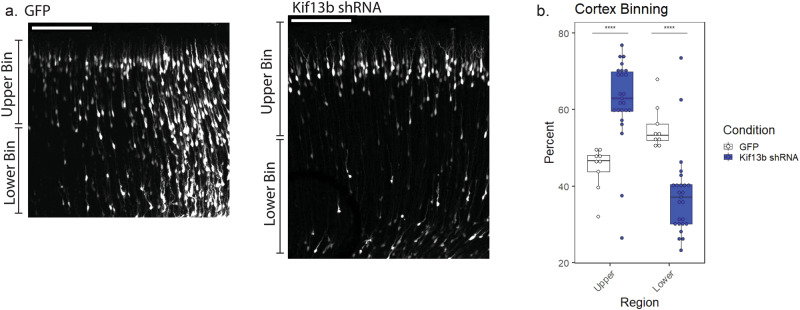
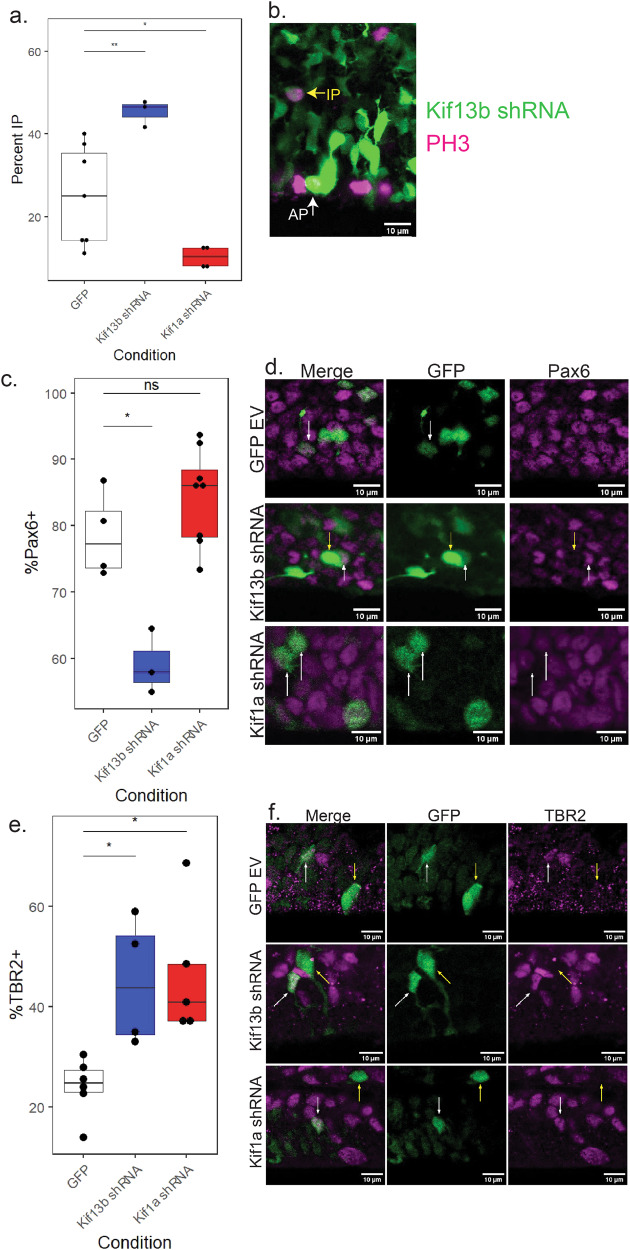
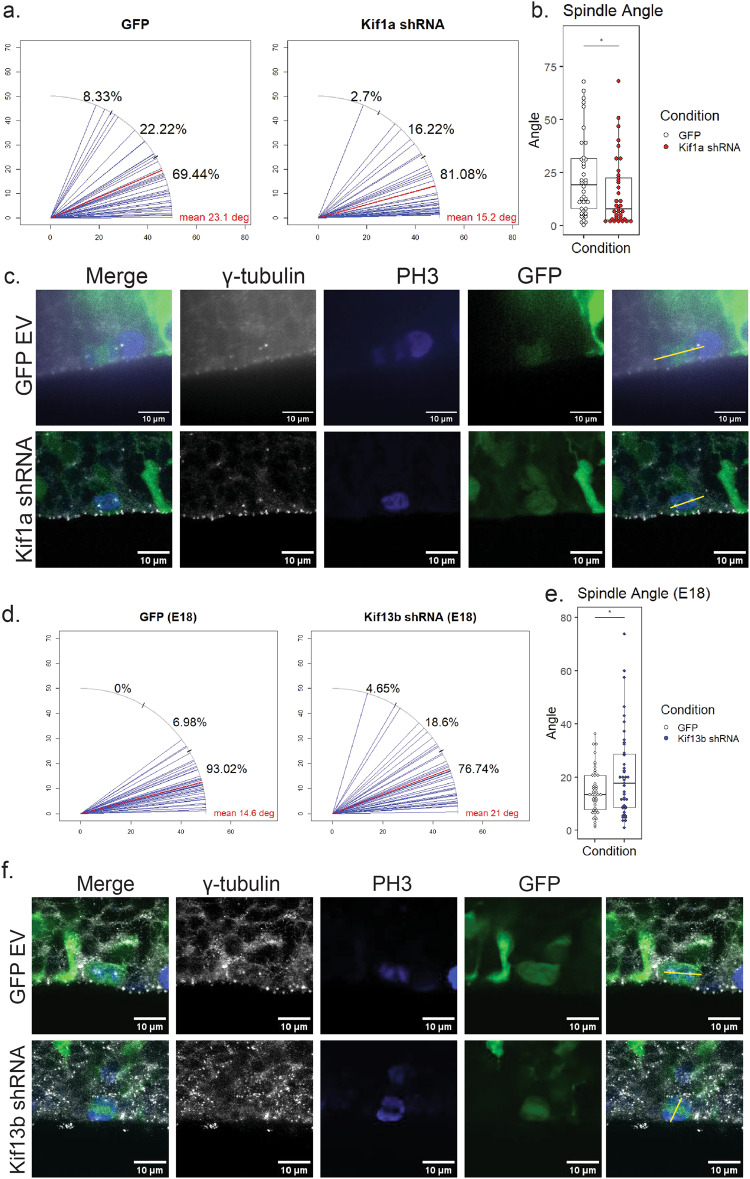
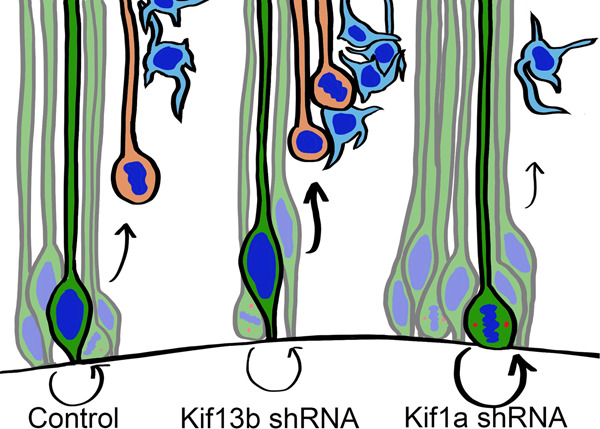
During the course of brain development, Radial Glial Progenitor (RGP) cells give rise to most of the neurons required for a functional cortex. RGPs can undergo symmetric divisions, which result in RGP duplication, or asymmetric divisions, which result in one RGP as well as one to four neurons. The control of this balance is not fully understood, but must be closely regulated to produce the cells required for a functioning cortex, and to maintain the stem cell pool. In this study, we show that the balance between symmetric and asymmetric RGP divisions is in part regulated by the actions of two kinesins, Kif1A and Kif13B, which we find have opposing roles in neurogenesis through their action on the mitotic spindle in dividing RGPs. We find that Kif1A promotes neurogenesis, whereas Kif13B promotes symmetric, non-neurogenic divisions. Interestingly, the two kinesins are closely related in structure, and members of the same kinesin-3 subfamily, thus their opposing effects on spindle orientation appear to represent a novel mechanism for the regulation of neurogenesis.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Shitamukai A, Konno D, Matsuzaki F. Oblique radial glial divisions in the developing mouse neocortex induce self-renewing progenitors outside the germinal zone that resemble primate outer subventricular zone progenitors. J. Neurosci. 2011;31:3683–3695. doi: 10.1523/JNEUROSCI.4773-10.2011. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
