

Insights into the ISG15 transfer cascade by the UBE1L activating enzyme
- PMID: 38042859
- PMCID: PMC10693564
- DOI: 10.1038/s41467-023-43711-3
Insights into the ISG15 transfer cascade by the UBE1L activating enzyme
Abstract
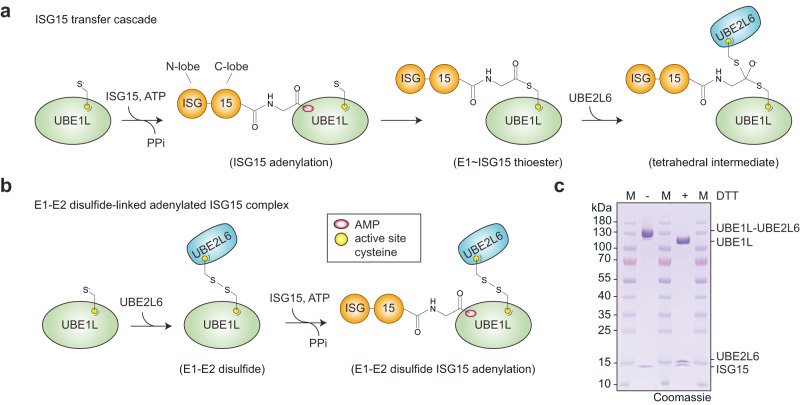
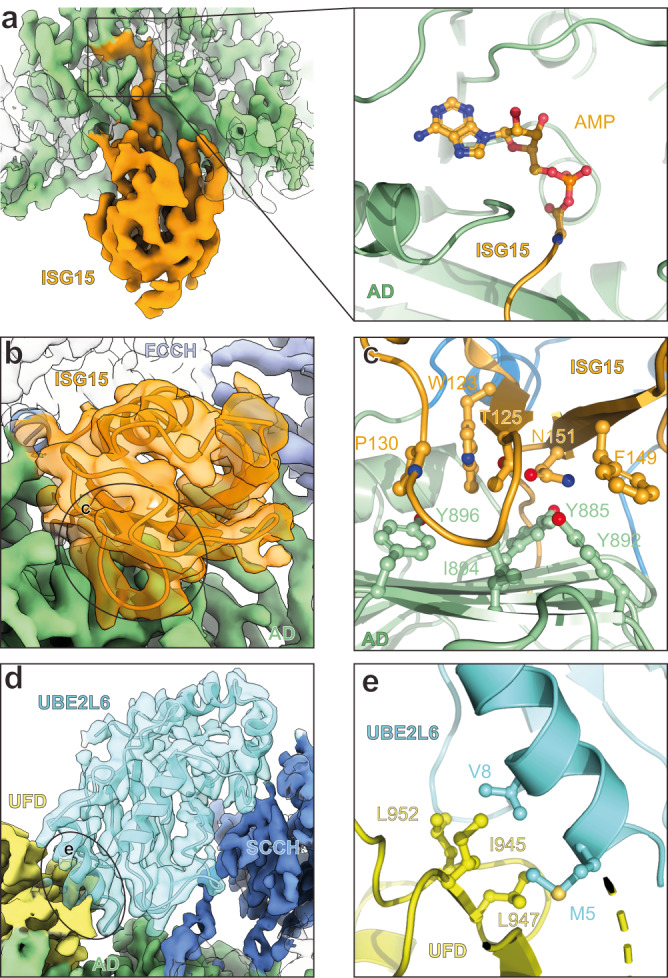
The attachment of the ubiquitin-like protein ISG15 to substrates by specific E1-E2-E3 enzymes is a well-established signalling mechanism of the innate immune response. Here, we present a 3.45 Å cryo-EM structure of a chemically trapped UBE1L-UBE2L6 complex bound to activated ISG15. This structure reveals the details of the first steps of ISG15 recognition and UBE2L6 recruitment by UBE1L (also known as UBA7). Taking advantage of viral effector proteins from severe acute respiratory coronavirus 2 (SARS-CoV-2) and influenza B virus (IBV), we validate the structure and confirm the importance of the ISG15 C-terminal ubiquitin-like domain in the adenylation reaction. Moreover, biochemical characterization of the UBE1L-ISG15 and UBE1L-UBE2L6 interactions enables the design of ISG15 and UBE2L6 mutants with altered selectively for the ISG15 and ubiquitin conjugation pathways. Together, our study helps to define the molecular basis of these interactions and the specificity determinants that ensure the fidelity of ISG15 signalling during the antiviral response.
© 2023. The Author(s).
Conflict of interest statement
B.A.S. is a member of the scientific advisory boards of Interline Therapeutics and BioTheryX. The remaining authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
- MC_UU_00018/10/RCUK | Medical Research Council (MRC)
- RGS_R2_222185/Royal Society
- 201302640/Deutsche Forschungsgemeinschaft (German Research Foundation)
- Nedd8Activate/EC | EU Framework Programme for Research and Innovation H2020 | H2020 Priority Excellent Science | H2020 European Research Council (H2020 Excellent Science - European Research Council)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
