

Macrophage LMO7 deficiency facilitates inflammatory injury via metabolic-epigenetic reprogramming
- PMID: 38045056
- PMCID: PMC10692378
- DOI: 10.1016/j.apsb.2023.09.012
Macrophage LMO7 deficiency facilitates inflammatory injury via metabolic-epigenetic reprogramming
Abstract
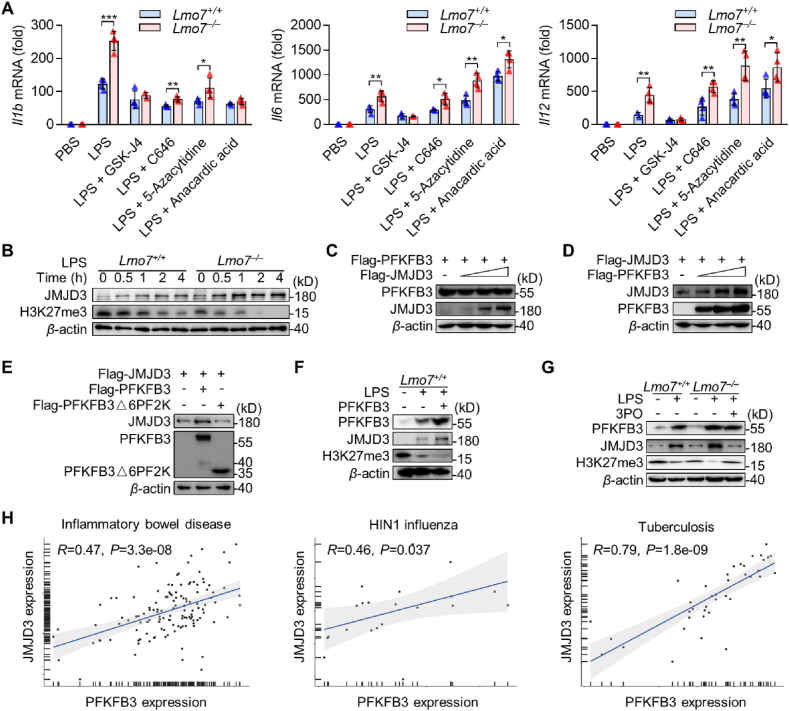
Inflammatory bowel disease (IBD) is a formidable disease due to its complex pathogenesis. Macrophages, as a major immune cell population in IBD, are crucial for gut homeostasis. However, it is still unveiled how macrophages modulate IBD. Here, we found that LIM domain only 7 (LMO7) was downregulated in pro-inflammatory macrophages, and that LMO7 directly degraded 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 (PFKFB3) through K48-mediated ubiquitination in macrophages. As an enzyme that regulates glycolysis, PFKFB3 degradation led to the glycolytic process inhibition in macrophages, which in turn inhibited macrophage activation and ultimately attenuated murine colitis. Moreover, we demonstrated that PFKFB3 was required for histone demethylase Jumonji domain-containing protein 3 (JMJD3) expression, thereby inhibiting the protein level of trimethylation of histone H3 on lysine 27 (H3K27me3). Overall, our results indicated the LMO7/PFKFB3/JMJD3 axis is essential for modulating macrophage function and IBD pathogenesis. Targeting LMO7 or macrophage metabolism could potentially be an effective strategy for treating inflammatory diseases.
Keywords: Inflammatory bowel disease; JMJD3; LMO7; Macrophage; PFKFB3; Ubiquitination.
© 2023 Chinese Pharmaceutical Association and Institute of Materia Medica, Chinese Academy of Medical Sciences. Production and hosting by Elsevier B.V.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures








References
-
- Bisgaard T.H., Allin K.H., Keefer L., Ananthakrishnan A.N., Jess T. Depression and anxiety in inflammatory bowel disease: epidemiology, mechanisms and treatment. Nat Rev Gastroenterol Hepatol. 2022;19:717–726. - PubMed
-
- Hegarty L.M., Jones G.R., Bain C.C. Macrophages in intestinal homeostasis and inflammatory bowel disease. Nat Rev Gastroenterol Hepatol. 2023;20:538–553. - PubMed
-
- Delfini M., Stakenborg N., Viola M.F., Boeckxstaens G. Macrophages in the gut: masters in multitasking. Immunity. 2022;55:1530–1548. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
