

This is a preprint.
Two-dose "extended priming" immunization amplifies humoral immune responses by synchronizing vaccine delivery with the germinal center response
- PMID: 38045401
- PMCID: PMC10690148
- DOI: 10.1101/2023.11.20.563479
Two-dose "extended priming" immunization amplifies humoral immune responses by synchronizing vaccine delivery with the germinal center response
Update in
-
Two-dose priming immunization amplifies humoral immunity by synchronizing vaccine delivery with the germinal center response.Sci Immunol. 2024 Sep 20;9(99):eadl3755. doi: 10.1126/sciimmunol.adl3755. Epub 2024 Sep 20. Sci Immunol. 2024. PMID: 39303017 Free PMC article.
Abstract
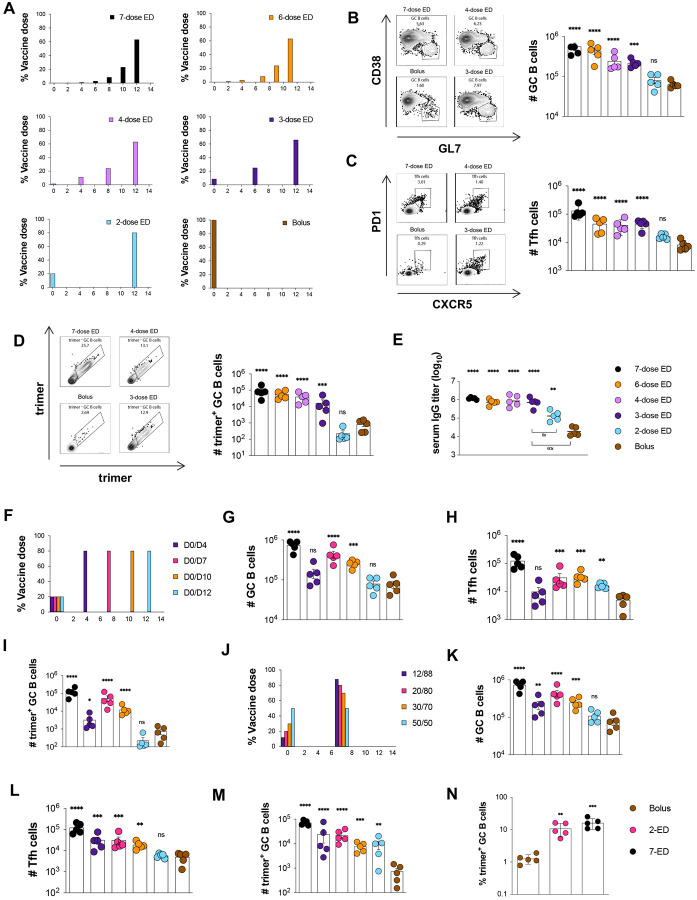
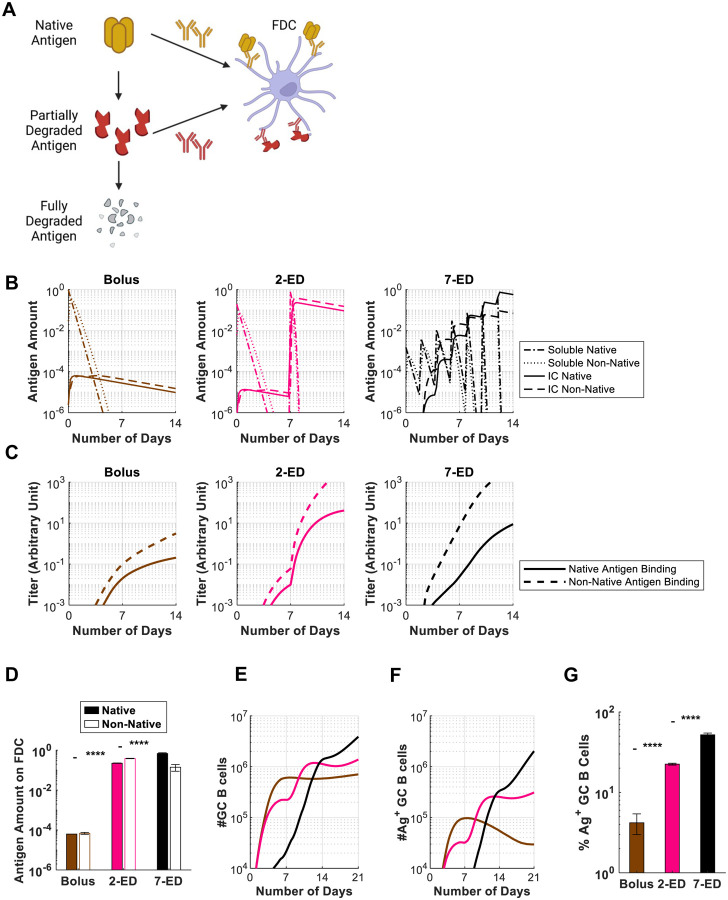
"Extended priming" immunization regimens that prolong exposure of the immune system to vaccines during the primary immune response have shown promise in enhancing humoral immune responses to a variety of subunit vaccines in preclinical models. We previously showed that escalating-dosing immunization (EDI), where a vaccine is dosed every other day in an increasing pattern over 2 weeks dramatically amplifies humoral immune responses. But such a dosing regimen is impractical for prophylactic vaccines. We hypothesized that simpler dosing regimens might replicate key elements of the immune response triggered by EDI. Here we explored "reduced ED" immunization regimens, assessing the impact of varying the number of injections, dose levels, and dosing intervals during EDI. Using a stabilized HIV Env trimer as a model antigen combined with a potent saponin adjuvant, we found that a two-shot extended-prime regimen consisting of immunization with 20% of a given vaccine dose followed by a second shot with the remaining 80% of the dose 7 days later resulted in increased total GC B cells, 5-10-fold increased frequencies of antigen-specific GC B cells, and 10-fold increases in serum antibody titers compared to single bolus immunization. Computational modeling of the GC response suggested that this enhanced response is mediated by antigen delivered in the second dose being captured more efficiently as immune complexes in follicles, predictions we verified experimentally. Our computational and experimental results also highlight how properly designed reduced ED protocols enhance activation and antigen loading of dendritic cells and activation of T helper cells to amplify humoral responses. These results suggest that a two-shot priming approach can be used to substantially enhance responses to subunit vaccines.
Conflict of interest statement
Competing interests S.H.B, D.J.I, A.K.C, and L.Y are inventors on a patent filing related to the extended-priming regimens described in this manuscript. For completeness, it is also noted that A.K.C. is a consultant (titled “Academic Partner”) for Flagship Pioneering, consultant and Strategic Oversight Board Member of its affiliated company, Apriori Bio, and is a consultant and Scientific Advisory Board Member of another affiliated company, Metaphore Bio.
Figures




References
-
- Feinberg M. B., Uhambo — Twists and Turns on the Journey to an Efficacious HIV-1 Vaccine. New Engl J Med. 384, 1157–1159 (2021). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous