

Genetic dissection of the degradation pathways for the mycotoxin fusaric acid in Burkholderia ambifaria T16
- PMID: 38054732
- PMCID: PMC10734416
- DOI: 10.1128/aem.00630-23
Genetic dissection of the degradation pathways for the mycotoxin fusaric acid in Burkholderia ambifaria T16
Abstract
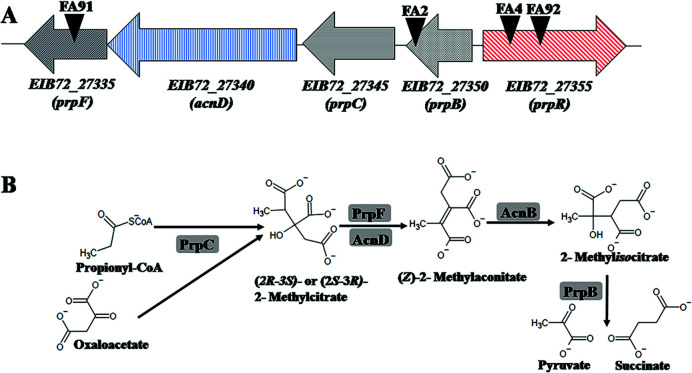
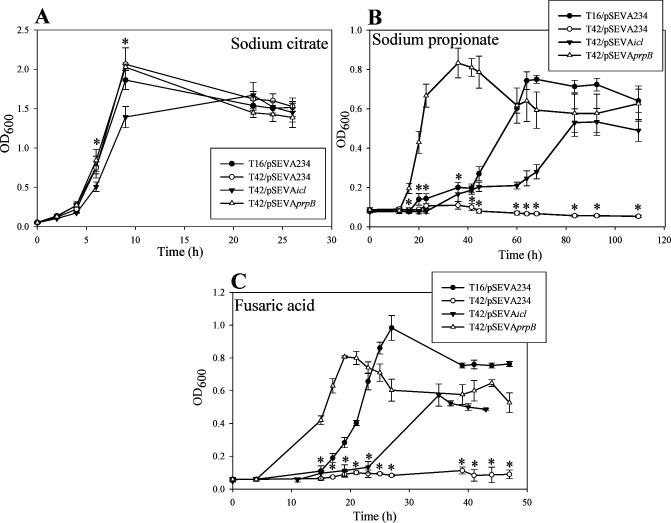
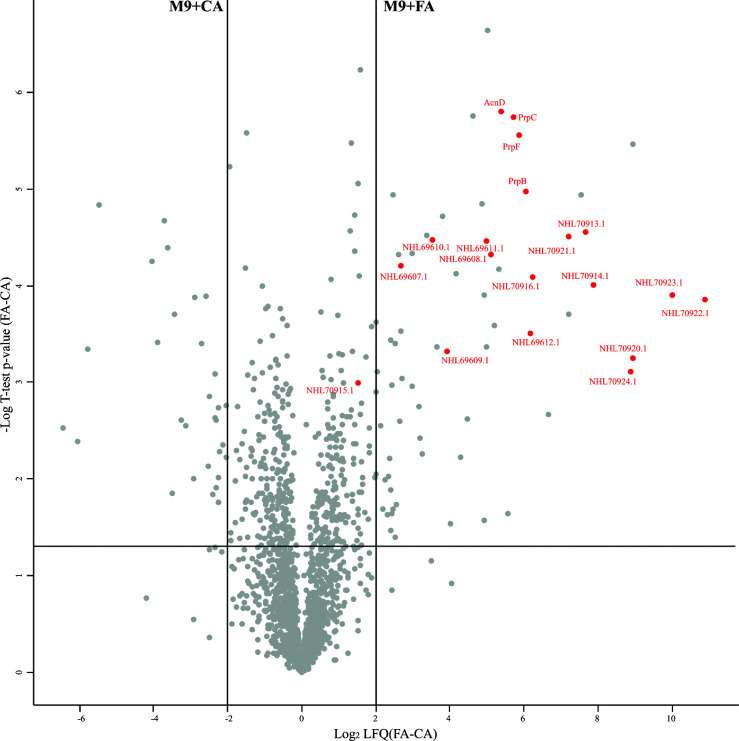
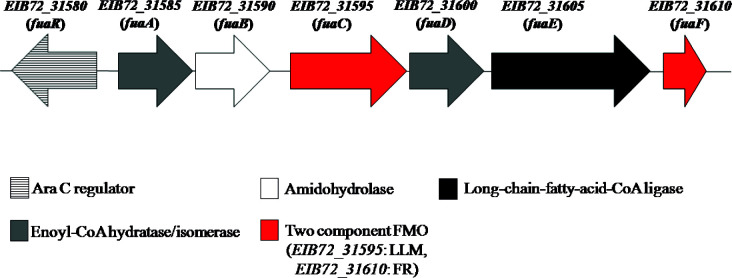
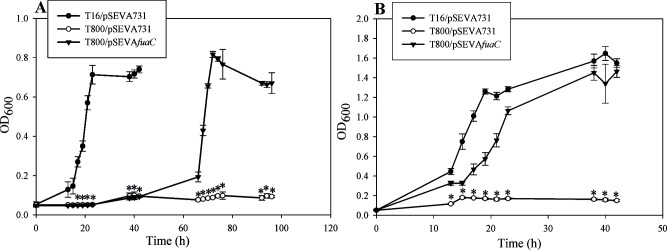
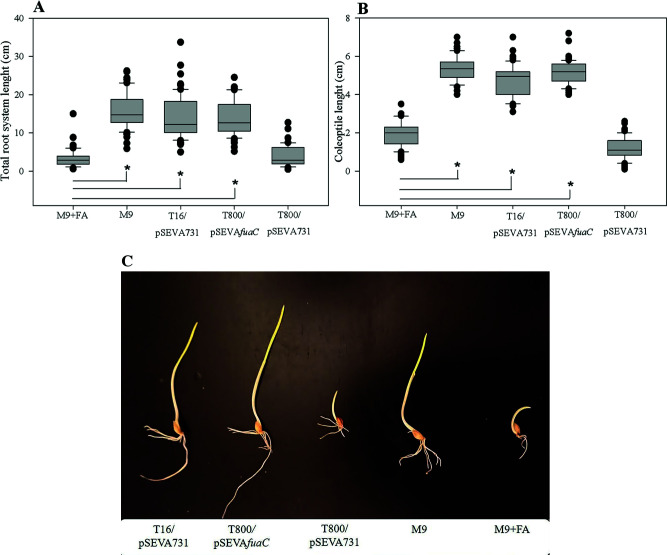
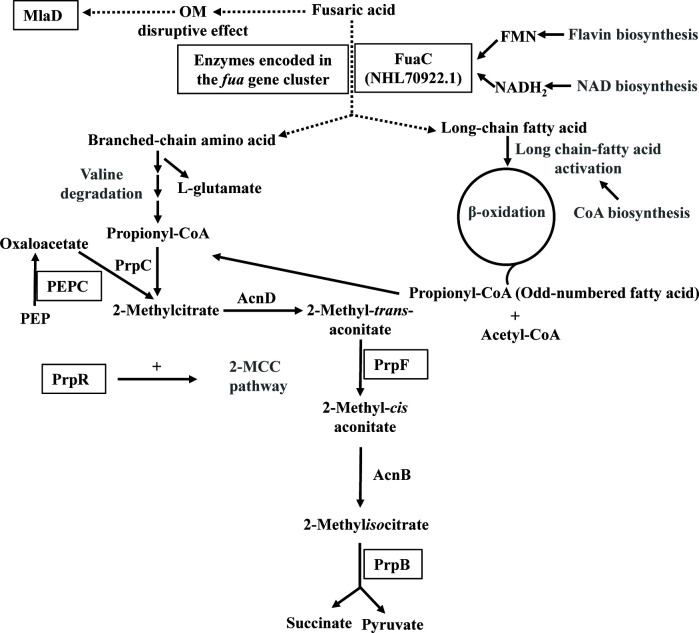
Fusaric acid (FA) is an important virulence factor produced by several Fusarium species. These fungi are responsible for wilt and rot diseases in a diverse range of crops. FA is toxic for animals, humans and soil-borne microorganisms. This mycotoxin reduces the survival and competition abilities of bacterial species able to antagonize Fusarium spp., due to its negative effects on viability and the production of antibiotics effective against these fungi. FA biodegradation is not a common characteristic among bacteria, and the determinants of FA catabolism have not been identified so far in any microorganism. In this study, we identified genes, enzymes, and metabolic pathways involved in the degradation of FA in the soil bacterium Burkholderia ambifaria T16. Our results provide insights into the catabolism of a pyridine-derivative involved in plant pathogenesis by a rhizosphere bacterium.
Keywords: 2-methylcitrate cycle; Burkholderia ambifaria T16; catabolism; detoxification; fusaric acid; two-component flavin-dependent monooxygenase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Pegg GF. 1981. Biochemistry and physiology of pathogenesis, p 193–246. In Mace ME, Bell AA, Beckman CH (ed), Fungal wilt diseases of plants. Academic Press Inc, New York.
-
- El-Hassan KI, El-Saman MG, Mosa AA, Mostafa M 2. 2007. Variation among Fusarium spp., the causal of potato tuber dry rot in their pathogenicity and mycotoxins production. Egypt J Phytopathol 35:53–68.
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- PICT-2017-1500/MINCyT | Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT)
- PICT-2020-SERIEA-02598/MINCyT | Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT)
- PUE0136/Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
- PIP0307/Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET)
- Alexander von Humboldt-Stiftung (AvH)
- NNF20CC0035580/Novo Nordisk Fonden (NNF)
- LiFe NNF18OC0034818/Novo Nordisk Fonden (NNF)
- TARGET NNF21OC0067996/Novo Nordisk Fonden (NNF)
- SWEET DFF-Research Project 8021-00039B/Danish Council for Independent Research
- No. 814418 SinFonia/EC | Horizon Europe | | HORIZON EUROPE European Innovation Council (EIC)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
