

Structural Dynamics of E6AP E3 Ligase HECT Domain and Involvement of a Flexible Hinge Loop in the Ubiquitin Chain Synthesis Mechanism
- PMID: 38055898
- PMCID: PMC10755755
- DOI: 10.1021/acs.nanolett.3c04150
Structural Dynamics of E6AP E3 Ligase HECT Domain and Involvement of a Flexible Hinge Loop in the Ubiquitin Chain Synthesis Mechanism
Abstract
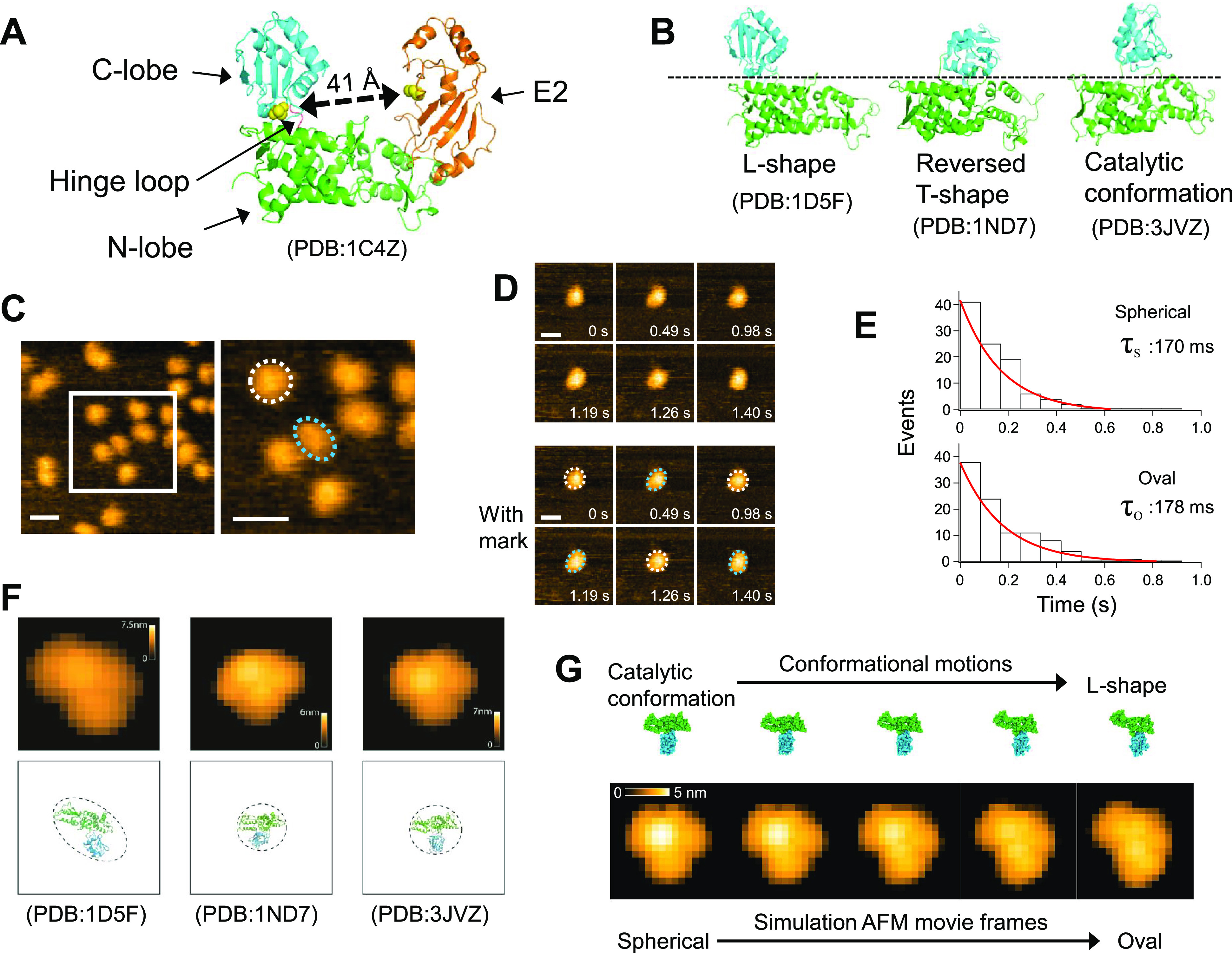
Ubiquitin (Ub) ligases E3 are important factors in selecting target proteins for ubiquitination and determining the type of polyubiquitin chains on the target proteins. In the HECT (homologous to E6AP C-terminus)-type E3 ligases, the HECT domain is composed of an N-lobe and a C-lobe that are connected by a flexible hinge loop. The large conformational rearrangement of the HECT domain via the flexible hinge loop is essential for the HECT-type E3-mediated Ub transfer from E2 to a target protein. However, detailed insights into the structural dynamics of the HECT domain remain unclear. Here, we provide the first direct demonstration of the structural dynamics of the HECT domain using high-speed atomic force microscopy at the nanoscale. We also found that the flexibility of the hinge loop has a great impact not only on its structural dynamics but also on the formation mechanism of free Ub chains.
Keywords: E6AP E3 ligase; HECT domain; high-speed atomic force microscope (HS-AFM); molecular modeling; structural dynamics; ubiquitin.
Conflict of interest statement
The authors declare no competing financial interest.
Figures




References
-
- Goldstein G.; Scheid M.; Hammerling U.; Schlesinger D. H.; Niall H. D.; Boyse E. A. Isolation of a polypeptide that has lymphocyte-differentiating properties and is probably represented universally in living cells. Proc. Natl. Acad. Sci. U. S. A. 1975, 72 (1), 11–5. 10.1073/pnas.72.1.11. - DOI - PMC - PubMed
-
- Kamadurai H. B.; Souphron J.; Scott D. C.; Duda D. M.; Miller D. J.; Stringer D.; Piper R. C.; Schulman B. A. Insights into ubiquitin transfer cascades from a structure of a UbcH5B approximately ubiquitin-HECT(NEDD4L) complex. Mol. Cell 2009, 36 (6), 1095–102. 10.1016/j.molcel.2009.11.010. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
