

Epinephrine inhibits PI3Kα via the Hippo kinases
- PMID: 38060450
- PMCID: PMC10809223
- DOI: 10.1016/j.celrep.2023.113535
Epinephrine inhibits PI3Kα via the Hippo kinases
Abstract
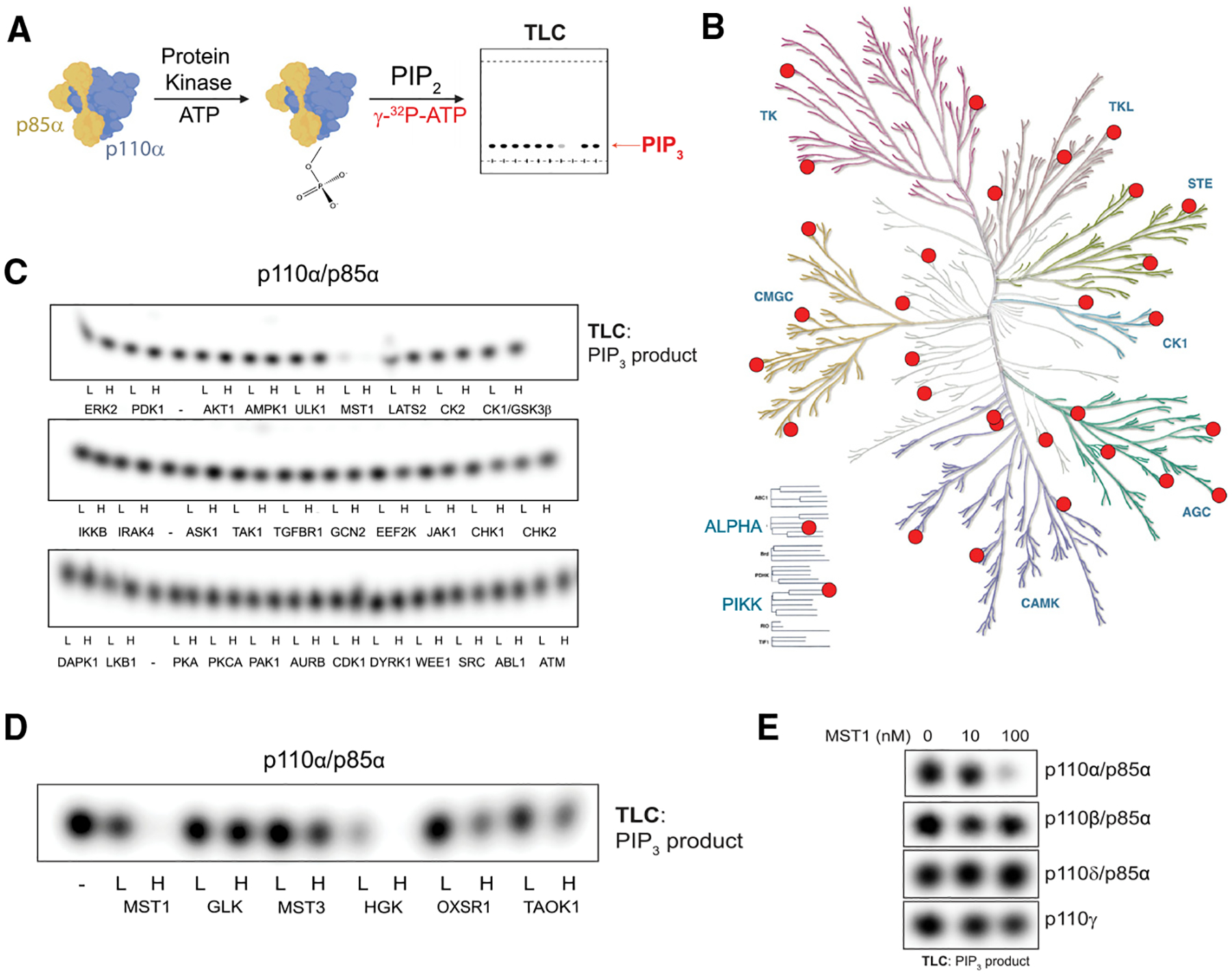
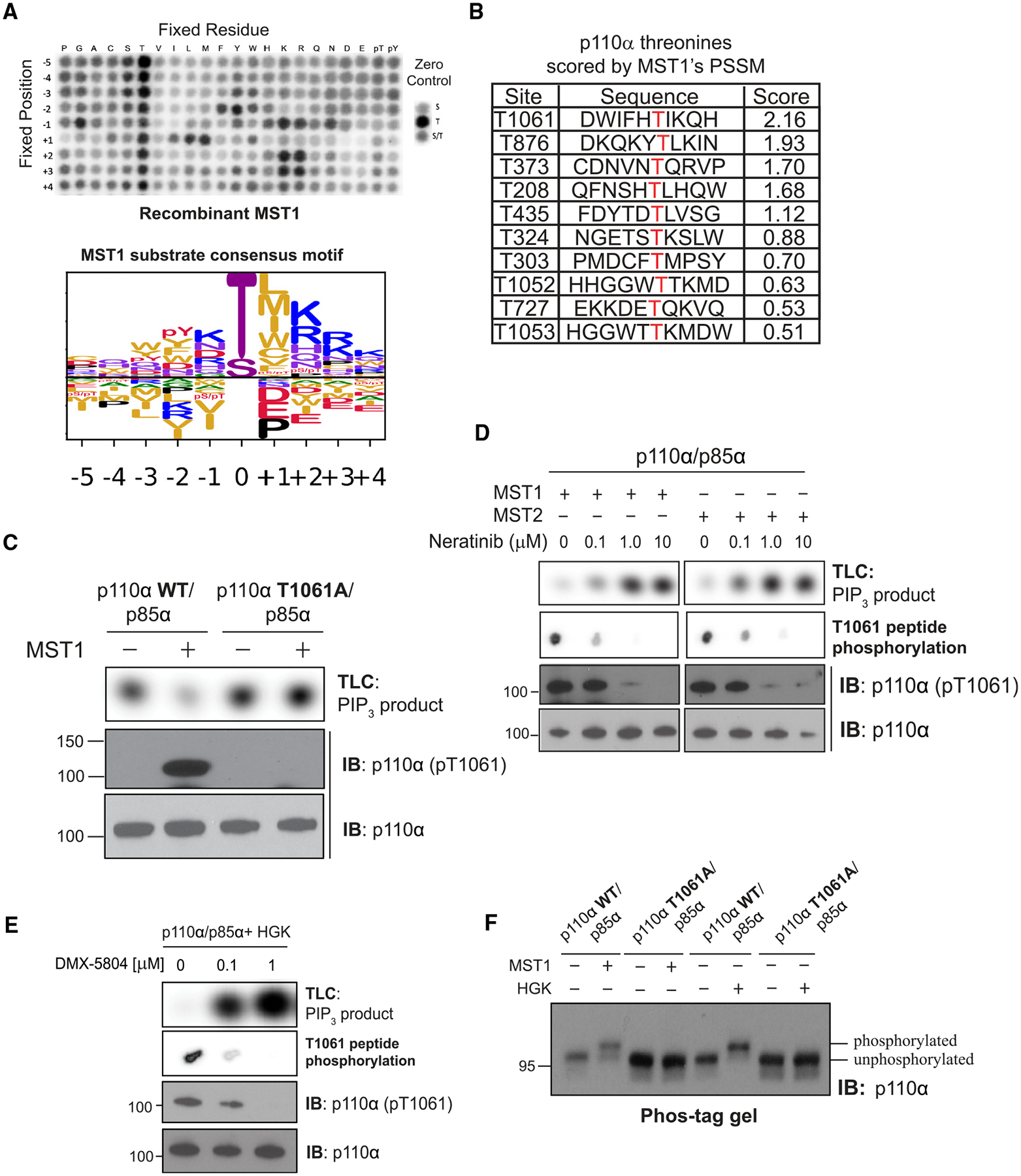
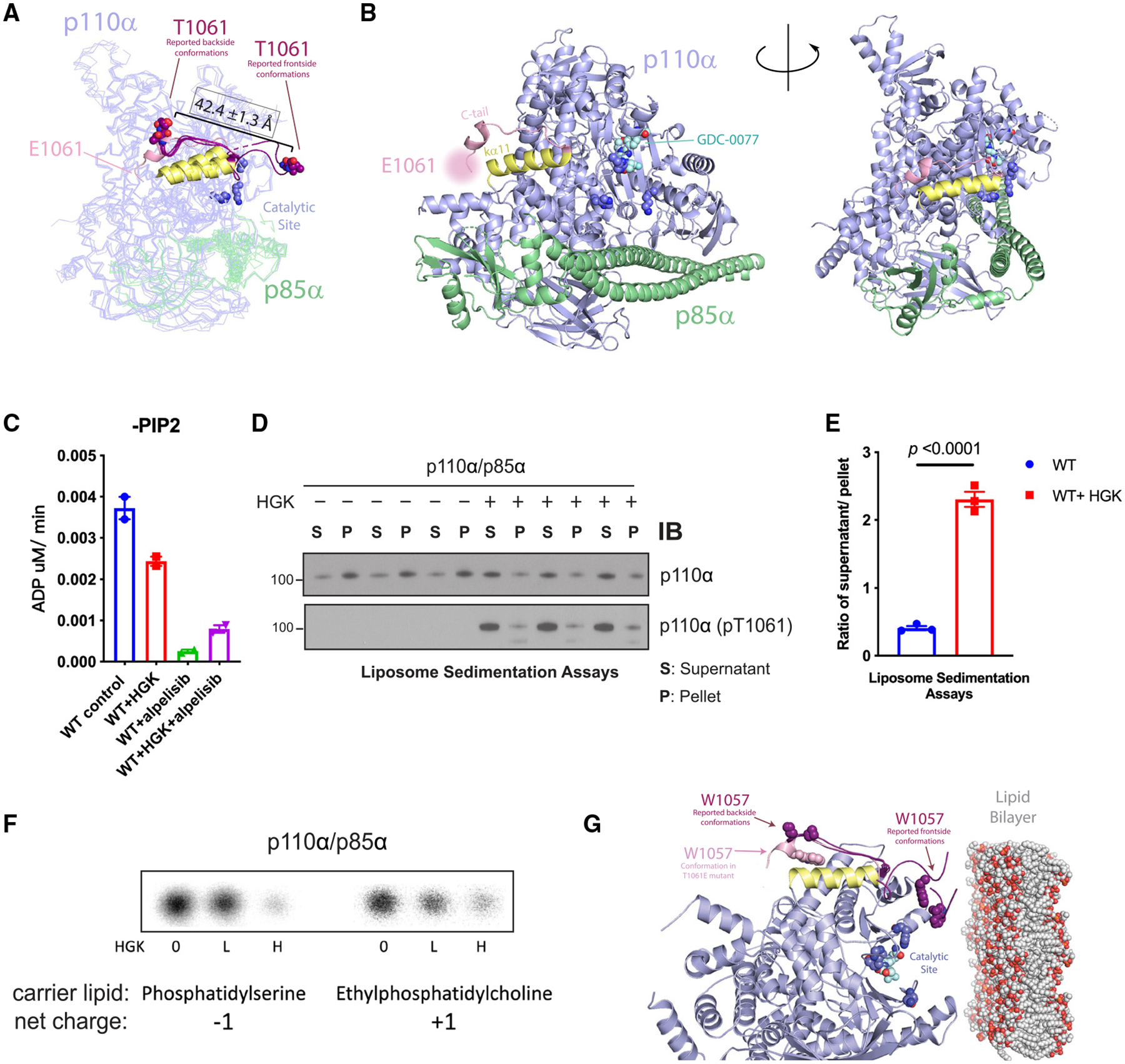
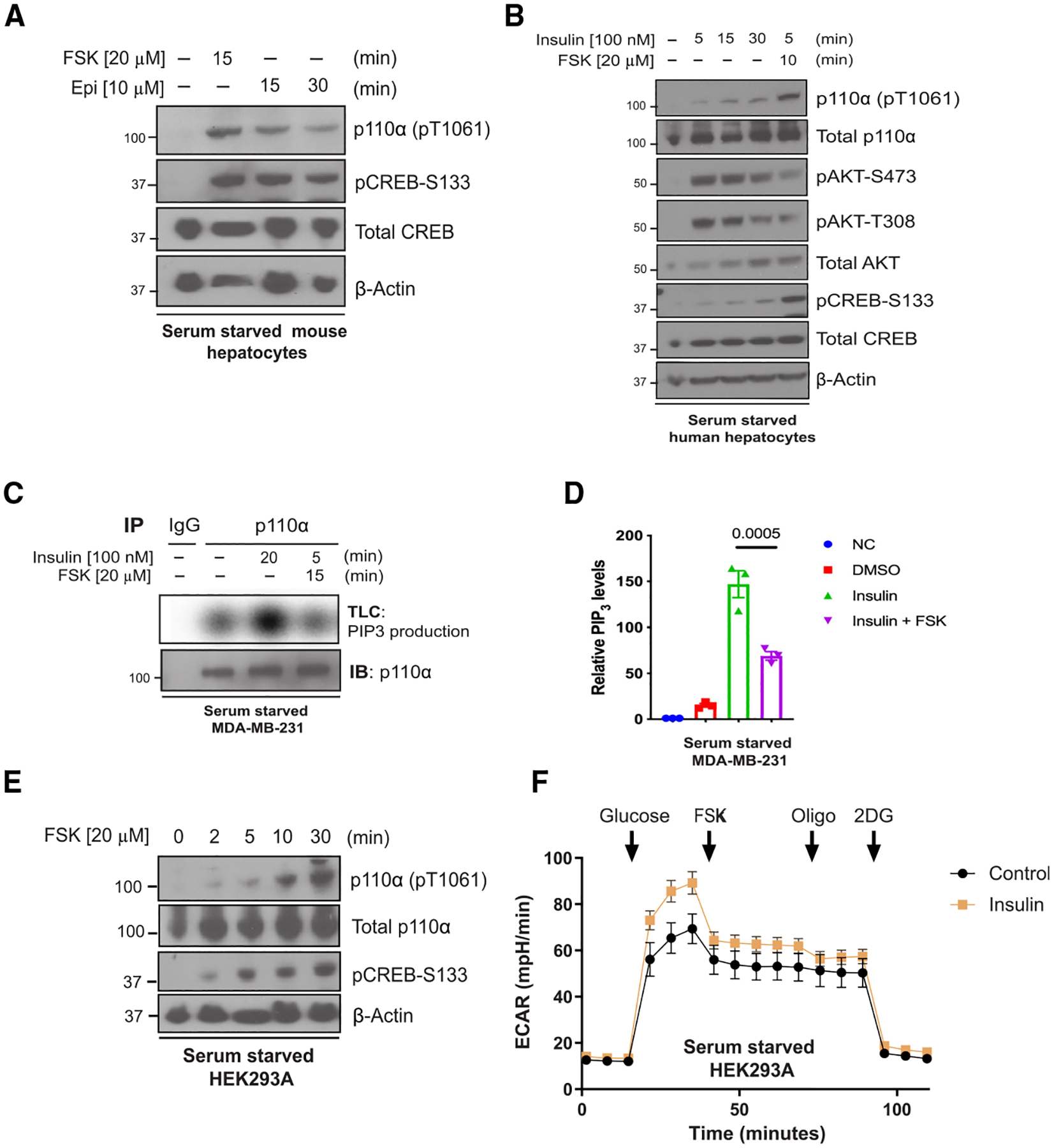
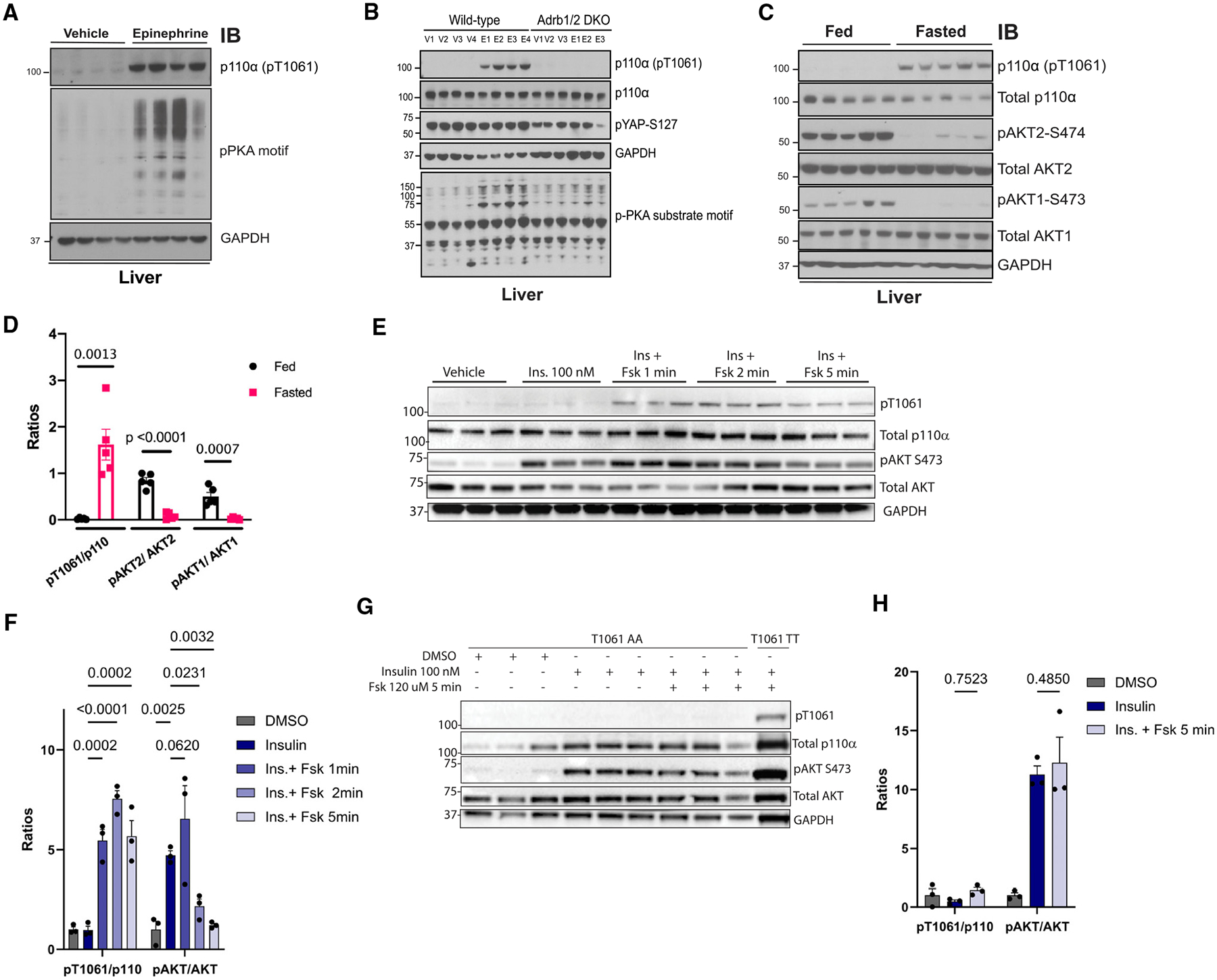
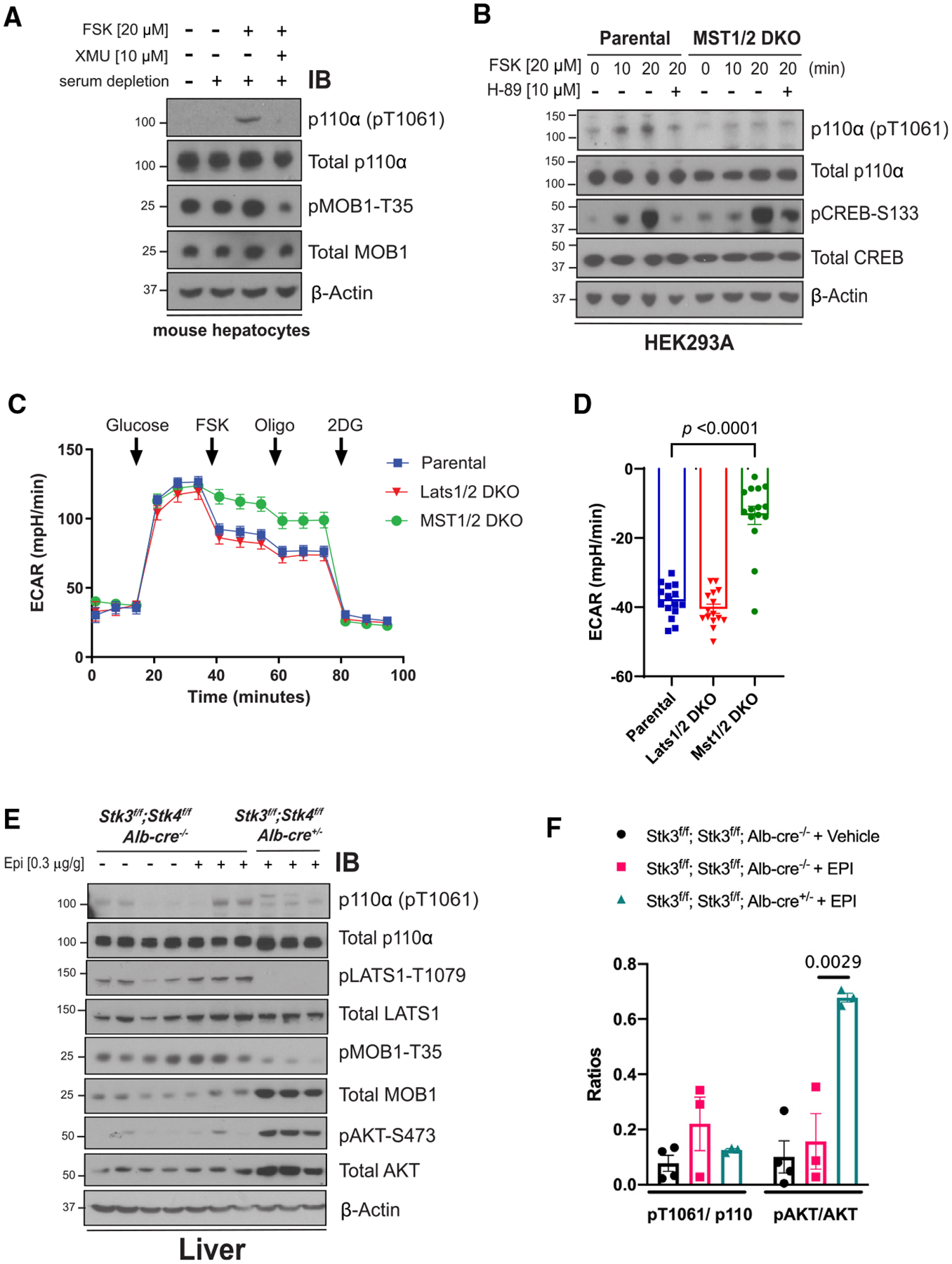
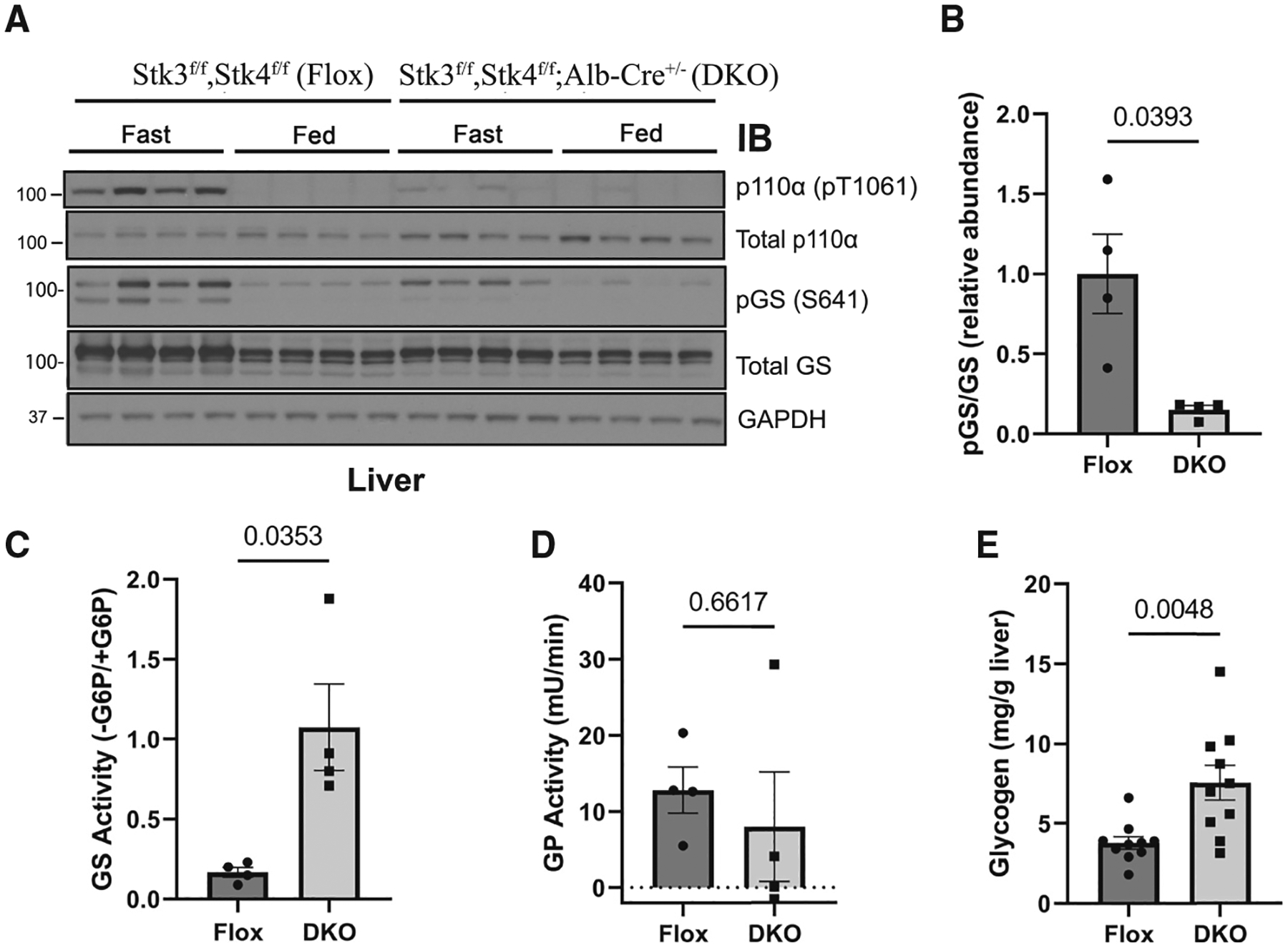
The phosphoinositide 3-kinase p110α is an essential mediator of insulin signaling and glucose homeostasis. We interrogated the human serine, threonine, and tyrosine kinome to search for novel regulators of p110α and found that the Hippo kinases phosphorylate p110α at T1061, which inhibits its activity. This inhibitory state corresponds to a conformational change of a membrane-binding domain on p110α, which impairs its ability to engage membranes. In human primary hepatocytes, cancer cell lines, and rodent tissues, activation of the Hippo kinases MST1/2 using forskolin or epinephrine is associated with phosphorylation of T1061 and inhibition of p110α, impairment of downstream insulin signaling, and suppression of glycolysis and glycogen synthesis. These changes are abrogated when MST1/2 are genetically deleted or inhibited with small molecules or if the T1061 is mutated to alanine. Our study defines an inhibitory pathway of PI3K signaling and a link between epinephrine and insulin signaling.
Keywords: CP: Metabolism; CP: Molecular biology; Hippo kinases; PI3K signaling; epinephrine signaling; glucose metabolism; glycogen metabolism; insulin sensitivity; liver.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests N.V. reports consultant and advisory board activities for Novartis, Petra Pharmaceuticals, Reactive Biosciences, and Magnet Biomedicine. E.A.K. is a shareholder of Eli Lilly and Company, and E.A.K. and K.K. are employees of Loxo Oncology at Lilly. L.C.C. is a founder and member of the board of directors of Agios Pharmaceuticals; is a founder and receives research support from Petra Pharmaceuticals; has equity in and consults for Cell Signaling Technologies, Volastra, Larkspur, and 1 Base Pharmaceuticals; and consults for Loxo-Lilly. J.L.J. has received consulting fees from Scorpion Therapeutics and Volastra Therapeutics. J.L.J. reports consultant activities for Scorpion Therapeutics and Volastra Therapeutics. M.D.G. reports personal fees from Novartis AG, Pfizer, Inc., and Scorpion Therapeutics. L.C.C., B.D.H., and M.D.G. are inventors on patents for Combination Therapy for PI3K-associated Disease or Disorder and The Identification of Therapeutic Interventions to Improve Response to PI3K Inhibitors for Cancer Treatment. B.D.H., L.C.C., and M.D.G. are co-founders and shareholders in Faeth Therapeutics. R.E.S. is on the sponsored advisory board for Miromatrix, Inc., and Lime Therapeutics and is a consultant and speaker for Alnylam, Inc. T.M.Y. is a stockholder and on the board of directors of DESTROKE, Inc., an early-stage start-up developing mobile technology for automated clinical stroke detection.
Figures
References
-
- Nolte RT, Eck MJ, Schlessinger J, Shoelson SE, and Harrison SC (1996). Crystal structure of the PI 3-kinase p85 amino-terminal SH2 domain and its phosphopeptide complexes. Nat. Struct. Biol 3, 364–374. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
