

Oleic acid availability impacts thymocyte preprogramming and subsequent peripheral Treg cell differentiation
- PMID: 38062135
- PMCID: PMC10918613
- DOI: 10.1038/s41590-023-01672-1
Oleic acid availability impacts thymocyte preprogramming and subsequent peripheral Treg cell differentiation
Abstract
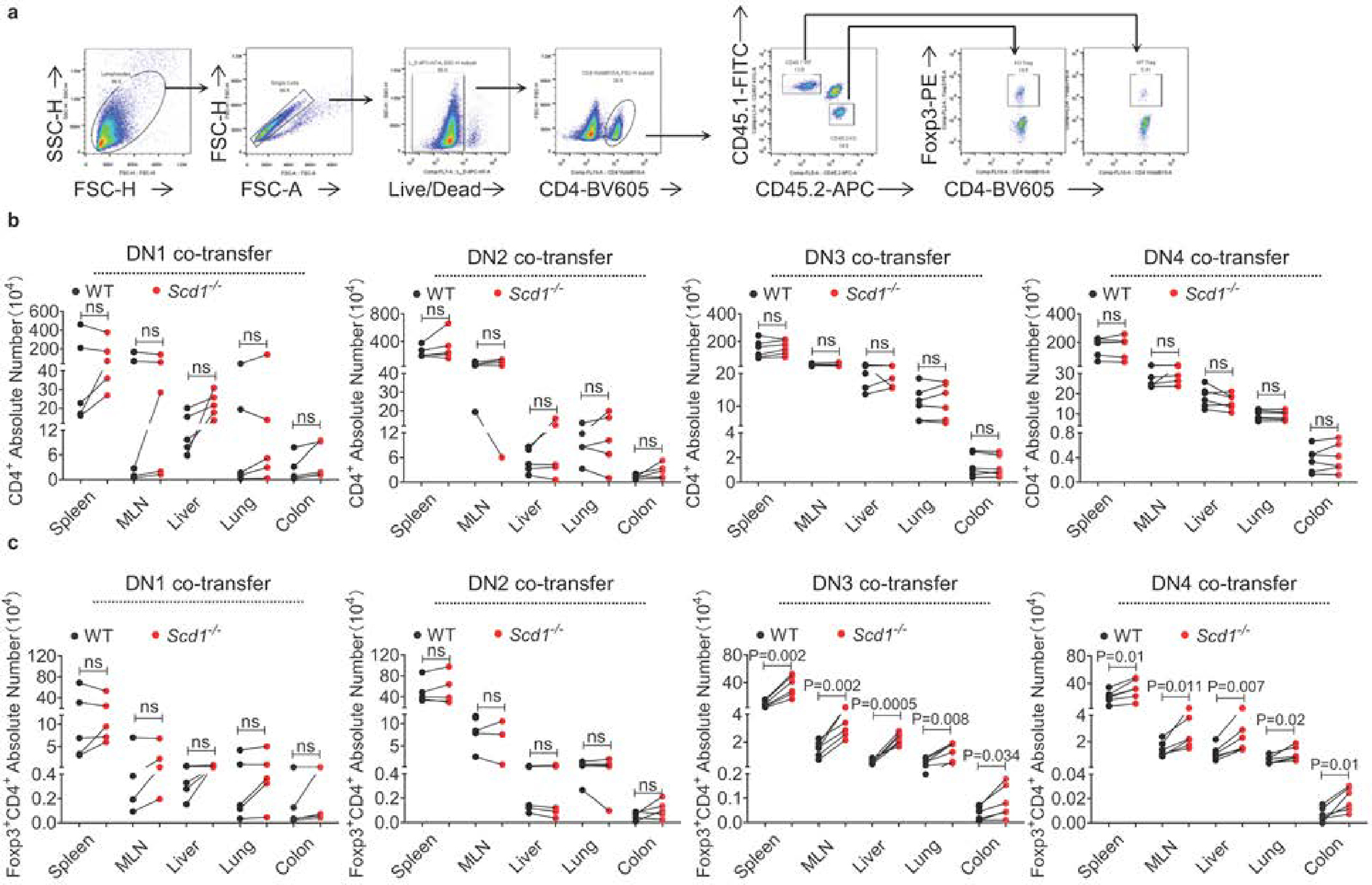
The nature of activation signals is essential in determining T cell subset differentiation; however, the features that determine T cell subset preference acquired during intrathymic development remain elusive. Here we show that naive CD4+ T cells generated in the mouse thymic microenvironment lacking Scd1, encoding the enzyme catalyzing oleic acid (OA) production, exhibit enhanced regulatory T (Treg) cell differentiation and attenuated development of experimental autoimmune encephalomyelitis. Scd1 deletion in K14+ thymic epithelia recapitulated the enhanced Treg cell differentiation phenotype of Scd1-deficient mice. The dearth of OA permitted DOT1L to increase H3K79me2 levels at the Atp2a2 locus of thymocytes at the DN2-DN3 transition stage. Such epigenetic modification persisted in naive CD4+ T cells and facilitated Atp2a2 expression. Upon T cell receptor activation, ATP2A2 enhanced the activity of the calcium-NFAT1-Foxp3 axis to promote naive CD4+ T cells to differentiate into Treg cells. Therefore, OA availability is critical for preprogramming thymocytes with Treg cell differentiation propensities in the periphery.
© 2023. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Ethics declarations
Competing interests
The authors declare no competing interests.
Figures
















References
-
- Cooper MD & Alder MN The evolution of adaptive immune systems. Cell 124, 815–822 (2006). - PubMed
-
- Irla M Instructive Cues of Thymic T Cell Selection. Annu Rev Immunol 40, 95–119 (2022). - PubMed
-
- Kadouri N, Nevo S, Goldfarb Y & Abramson J Thymic epithelial cell heterogeneity: TEC by TEC. Nat Rev Immunol 20, 239–253 (2020). - PubMed
-
- Taniuchi I CD4 Helper and CD8 Cytotoxic T Cell Differentiation. Annu Rev Immunol 36, 579–601 (2018). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
