

Antigen-specific memory NK cell responses against HIV and influenza use the NKG2/HLA-E axis
- PMID: 38064568
- PMCID: PMC11104516
- DOI: 10.1126/sciimmunol.adi3974
Antigen-specific memory NK cell responses against HIV and influenza use the NKG2/HLA-E axis
Abstract
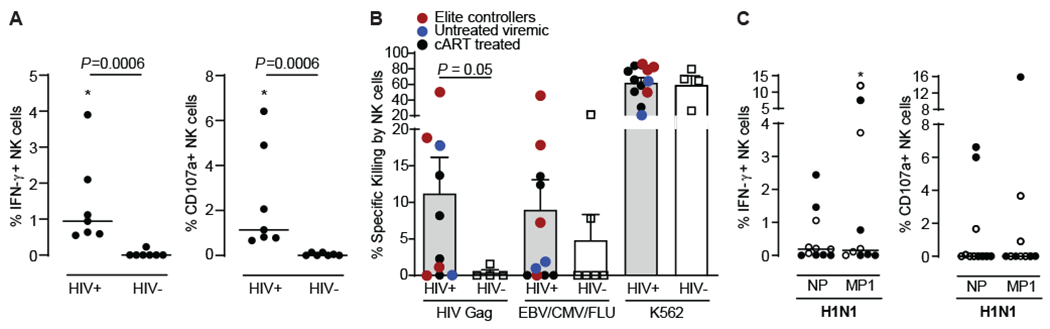
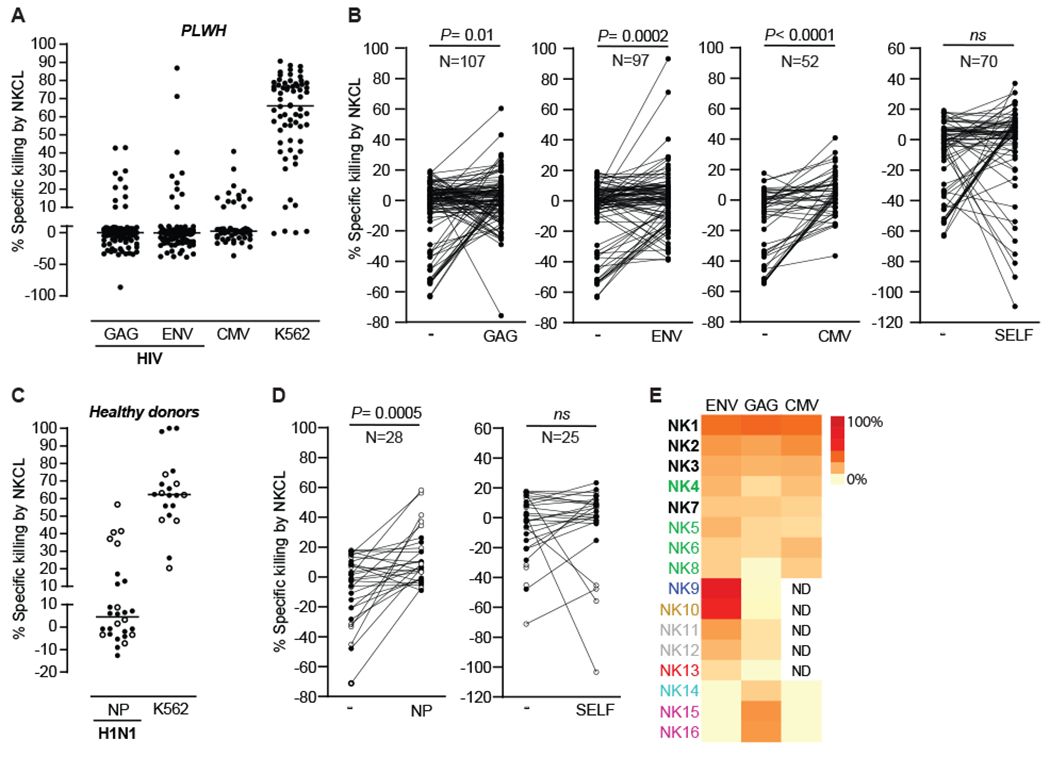
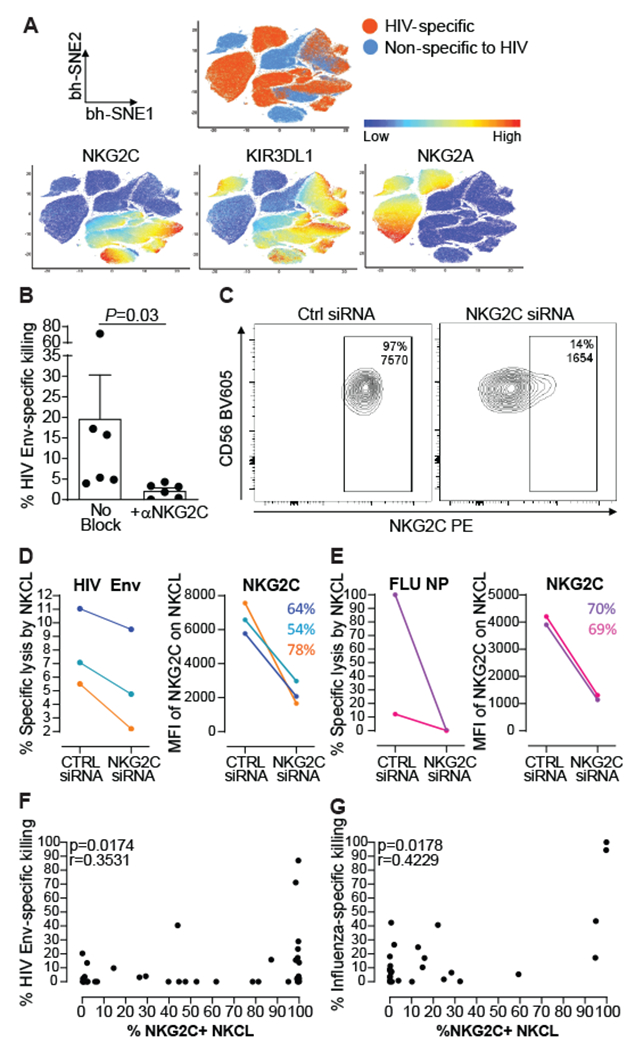
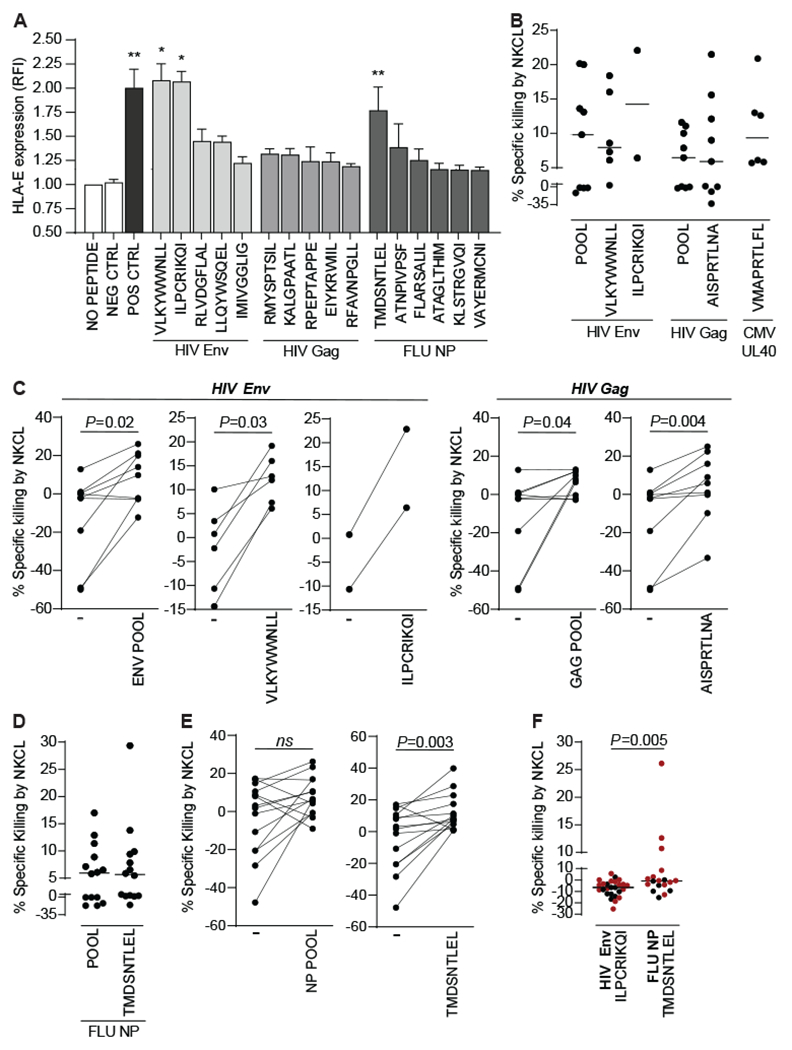
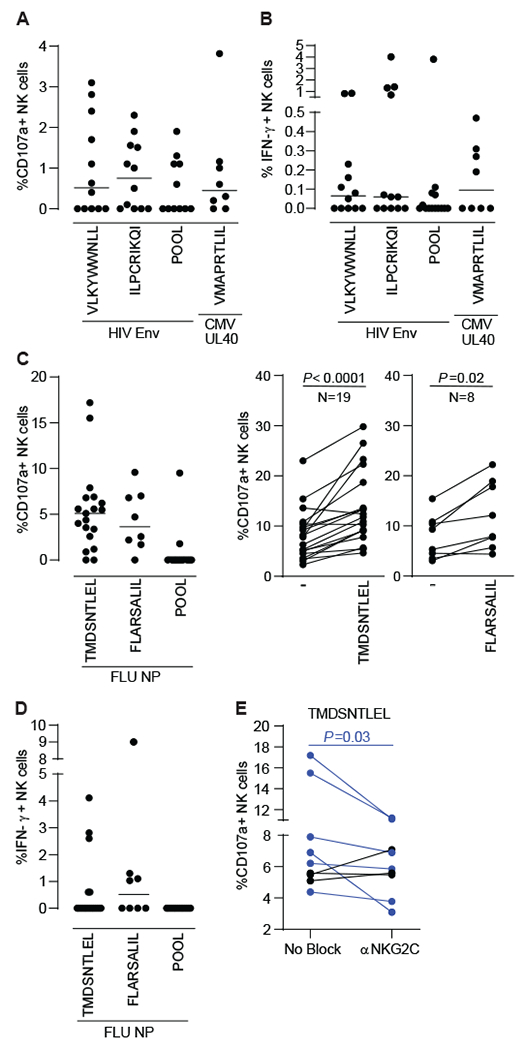
Multiple studies have broadened the roles of natural killer (NK) cells functioning as purely innate lymphocytes by demonstrating that they are capable of putative antigen-specific immunological memory against multiple infectious agents including HIV-1 and influenza. However, the mechanisms underlying antigen specificity remain unknown. Here, we demonstrate that antigen-specific human NK cell memory develops upon exposure to both HIV and influenza, unified by a conserved and epitope-specific targetable mechanism largely dependent on the activating CD94/NKG2C receptor and its ligand HLA-E. We validated the permanent acquisition of antigen specificity by individual memory NK cells by single-cell cloning. We identified elevated expression of KLRG1, α4β7, and NKG2C as biomarkers of antigen-specific NK cell memory through complex immunophenotyping. Last, we uncovered individual HLA-E-restricted peptides that may constitute the dominant NK cell response in HIV-1- and influenza-infected persons in vivo. Our findings clarify the mechanisms contributing to antigen-specific memory NK cell responses and suggest that they could be potentially targeted therapeutically for vaccines or other therapeutic interventions.
Conflict of interest statement
Competing Interests
The authors declare no competing financial interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
