

LncRNA MIR200CHG inhibits EMT in gastric cancer by stabilizing miR-200c from target-directed miRNA degradation
- PMID: 38065939
- PMCID: PMC10709323
- DOI: 10.1038/s41467-023-43974-w
LncRNA MIR200CHG inhibits EMT in gastric cancer by stabilizing miR-200c from target-directed miRNA degradation
Abstract
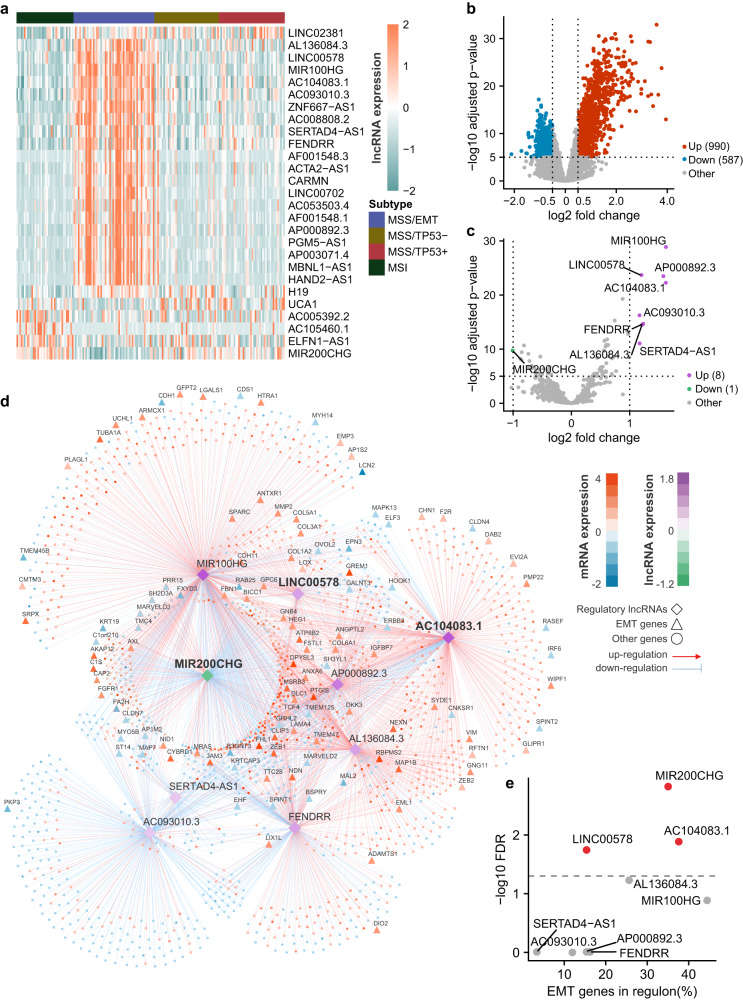
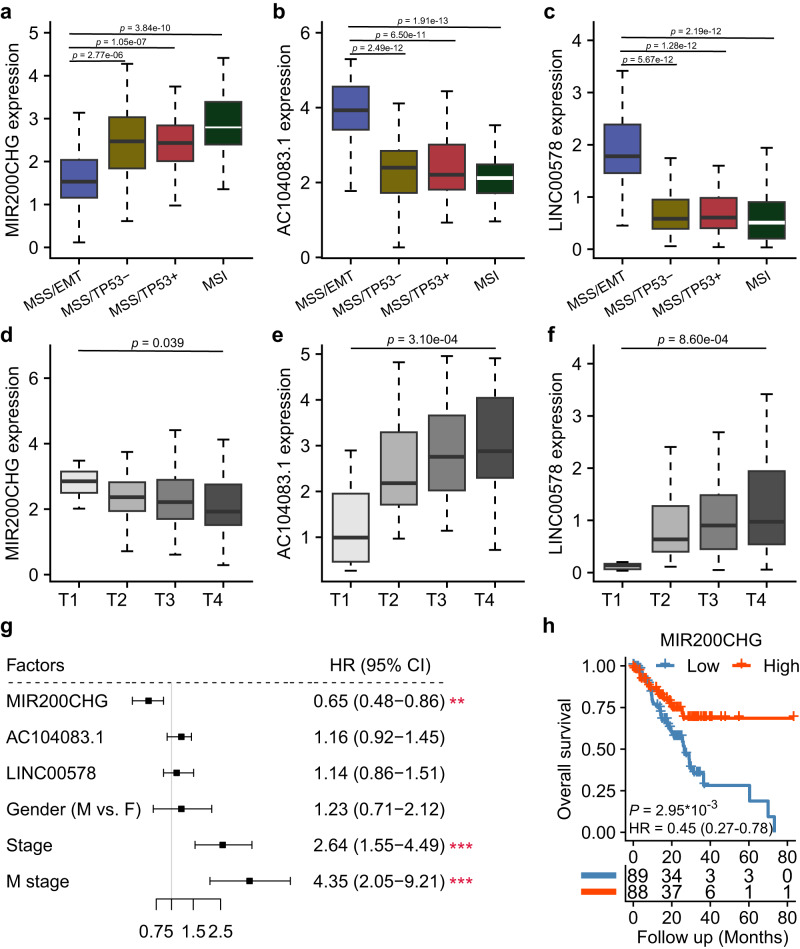
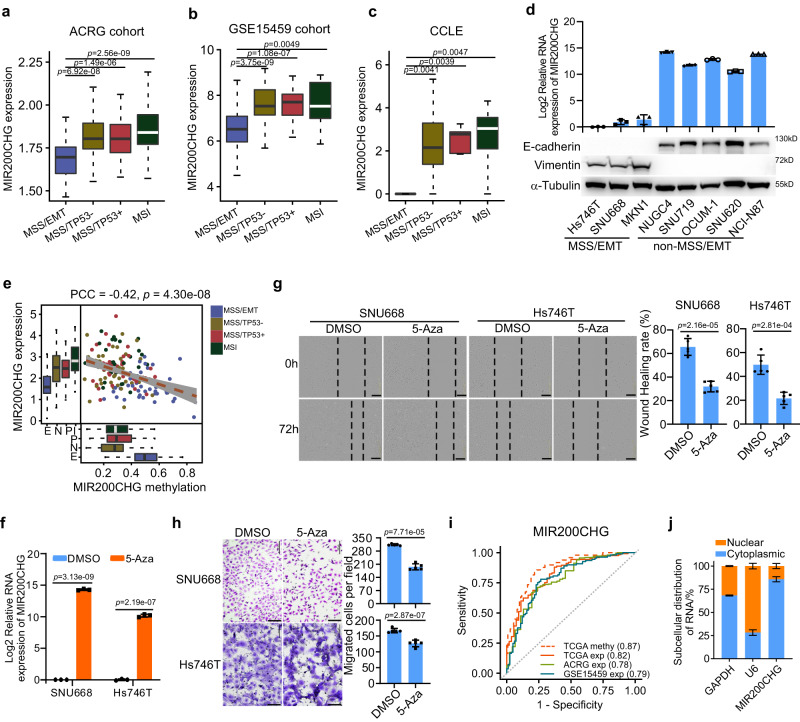
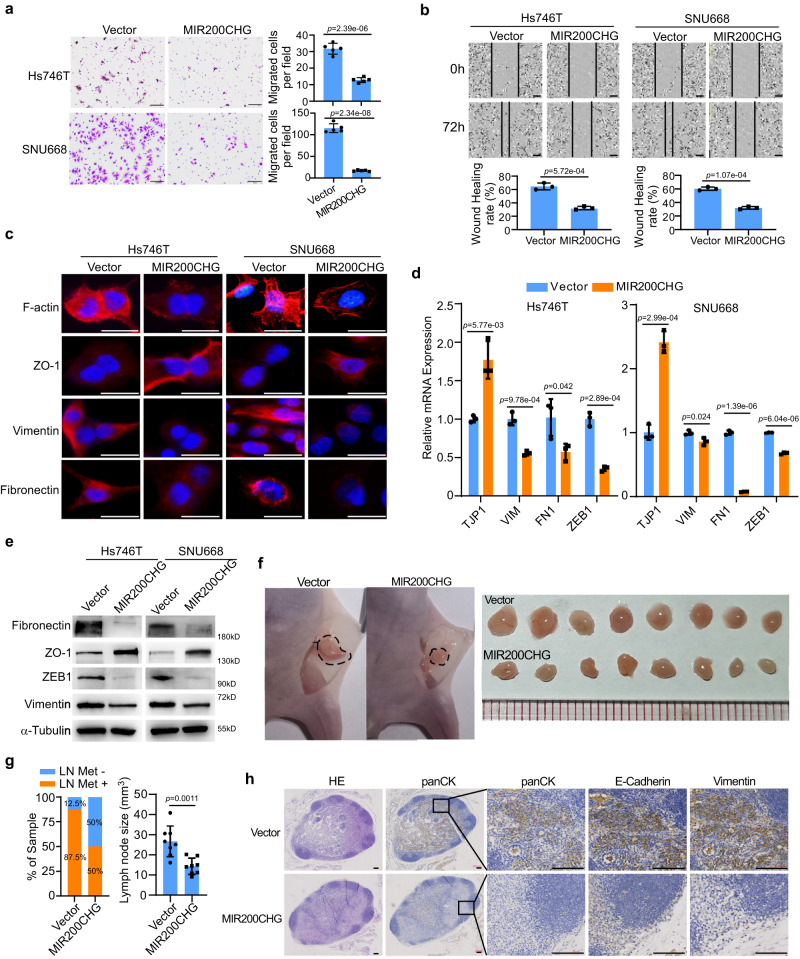
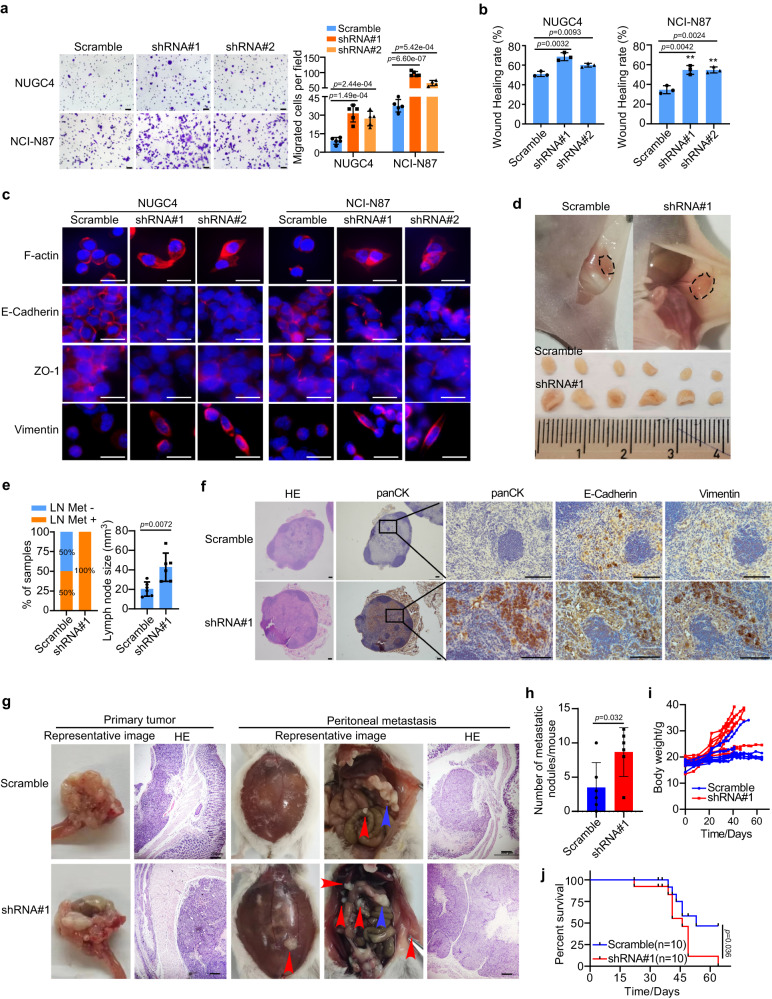
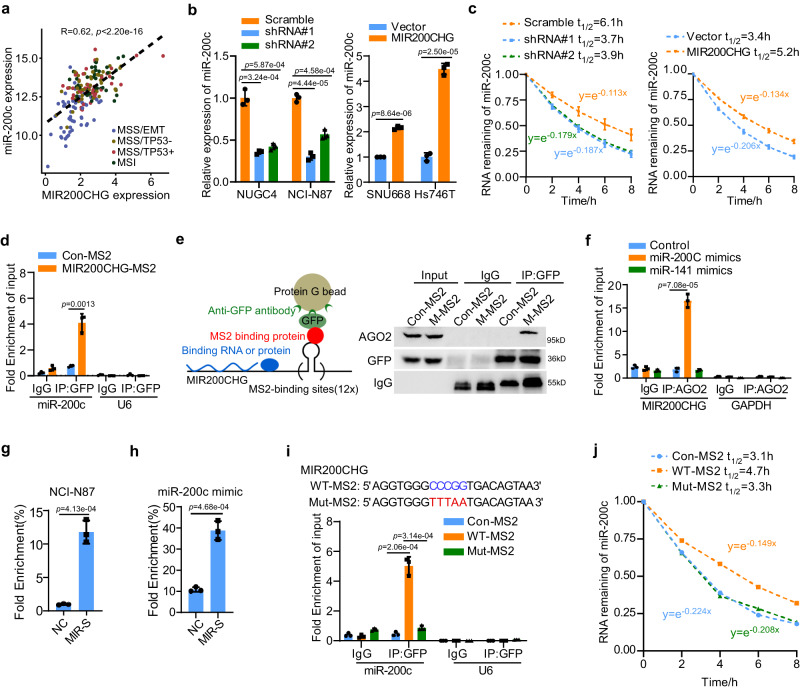
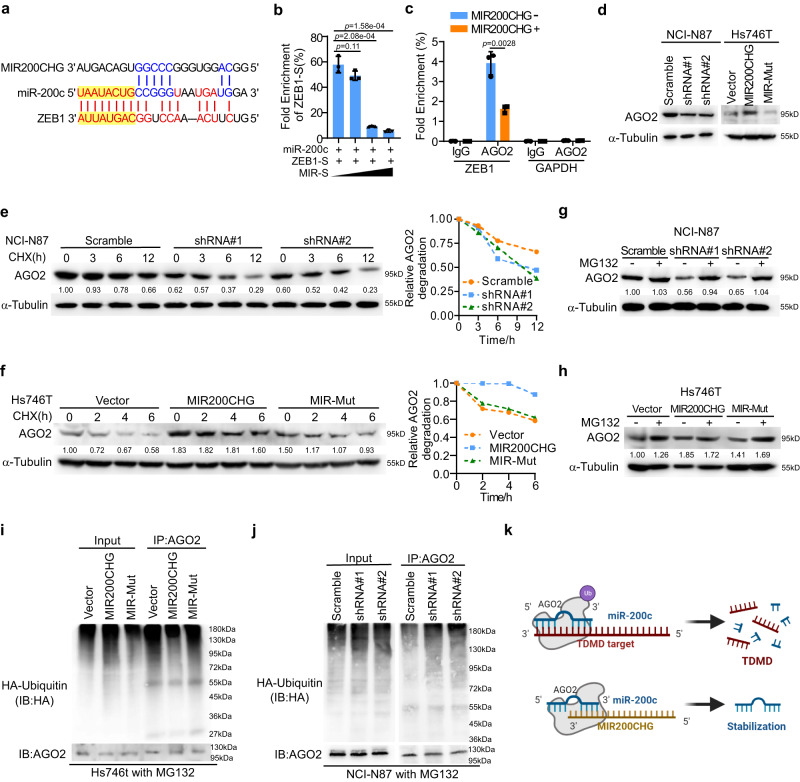
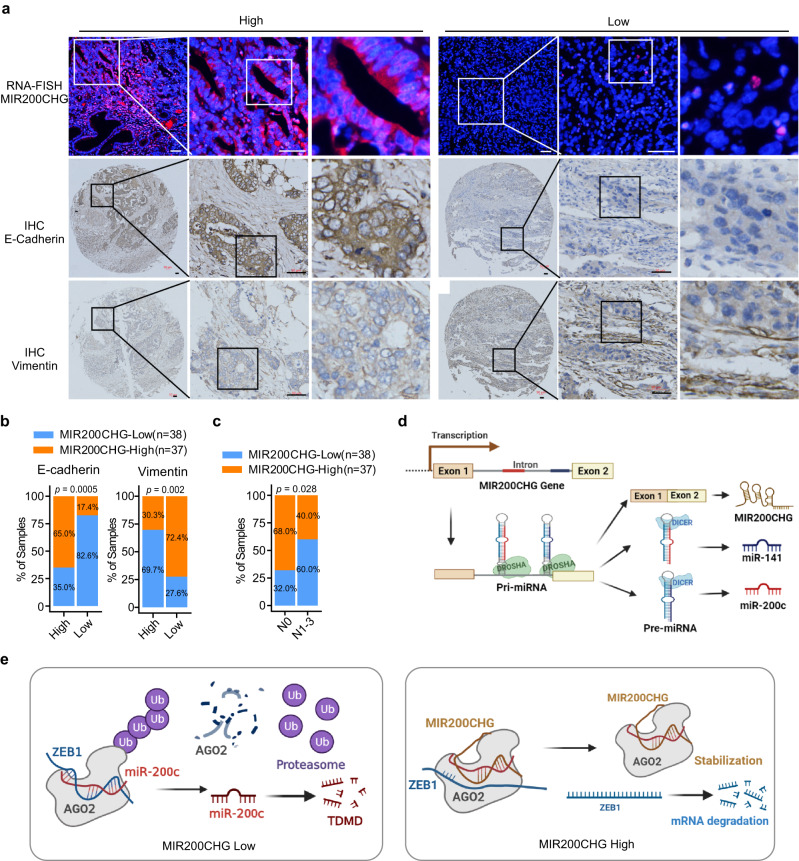
Gastric cancer (GC) is a heterogeneous disease, threatening millions of lives worldwide, yet the functional roles of long non-coding RNAs (lncRNAs) in different GC subtypes remain poorly characterized. Microsatellite stable (MSS)/epithelial-mesenchymal transition (EMT) GC is the most aggressive subtype associated with a poor prognosis. Here, we apply integrated network analysis to uncover lncRNA heterogeneity between GC subtypes, and identify MIR200CHG as a master regulator mediating EMT specifically in MSS/EMT GC. The expression of MIR200CHG is silenced in MSS/EMT GC by promoter hypermethylation, associated with poor prognosis. MIR200CHG reverses the mesenchymal identity of GC cells in vitro and inhibits metastasis in vivo. Mechanistically, MIR200CHG not only facilitates the biogenesis of its intronic miRNAs miR-200c and miR-141, but also protects miR-200c from target-directed miRNA degradation (TDMD) through direct binding to miR-200c. Our studies reveal a landscape of a subtype-specific lncRNA regulatory network, providing clinically relevant biological insights towards MSS/EMT GC.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
- 2020N368/Shenzhen Science and Technology Innovation Commission
- C4024-22GF/Research Grants Council, University Grants Committee (RGC, UGC)
- 14104223/Research Grants Council, University Grants Committee (RGC, UGC)
- 11103619/Research Grants Council, University Grants Committee (RGC, UGC)
- 14111522/Research Grants Council, University Grants Committee (RGC, UGC)
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
