

Targeting Metabolic Vulnerabilities to Overcome Prostate Cancer Resistance: Dual Therapy with Apalutamide and Complex I Inhibition
- PMID: 38067315
- PMCID: PMC10705744
- DOI: 10.3390/cancers15235612
Targeting Metabolic Vulnerabilities to Overcome Prostate Cancer Resistance: Dual Therapy with Apalutamide and Complex I Inhibition
Abstract
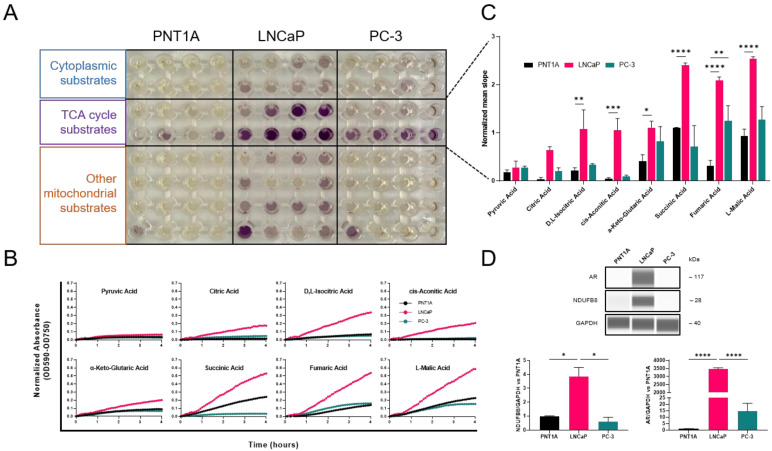
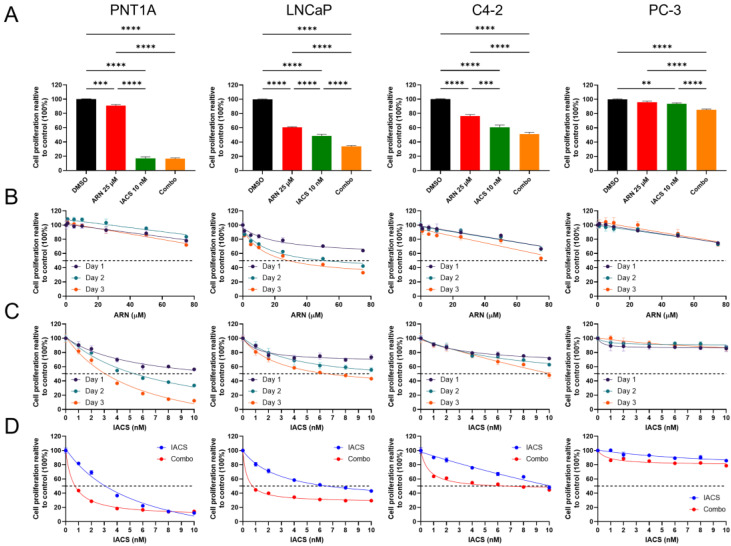
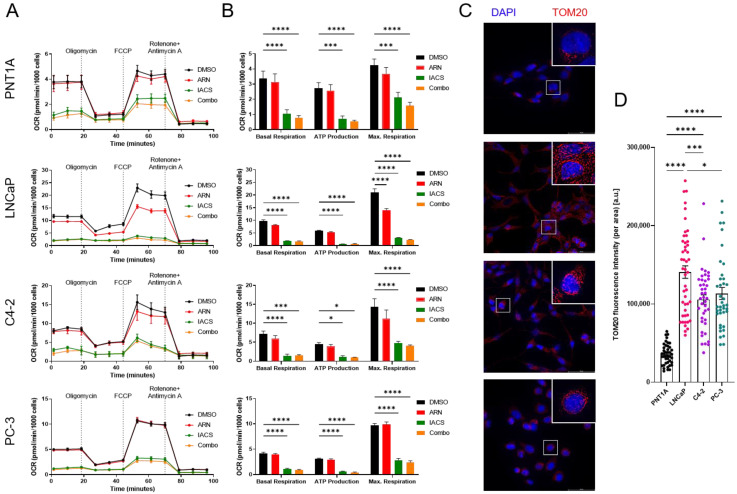
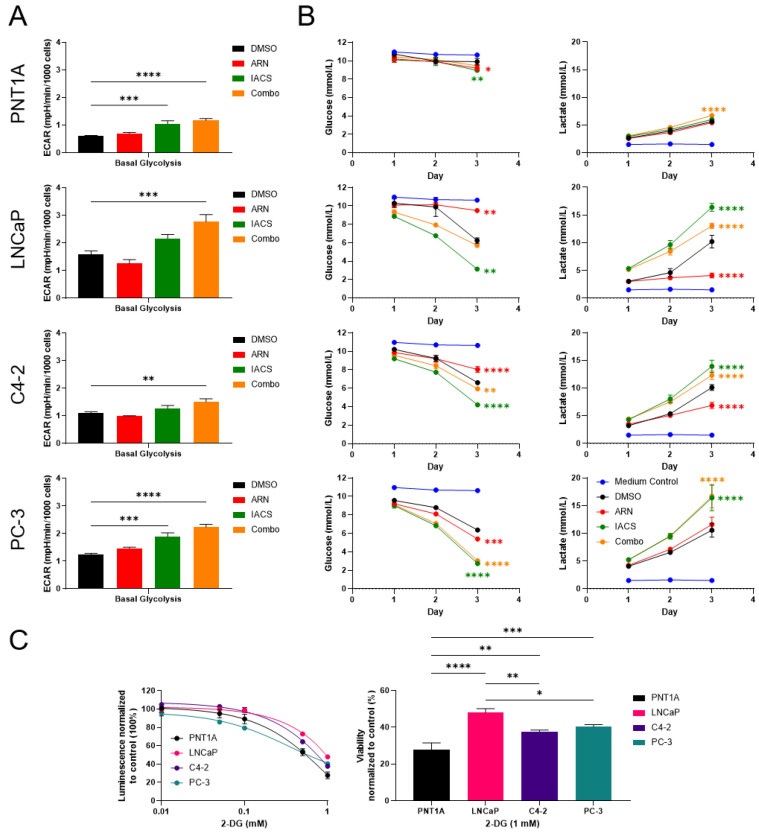
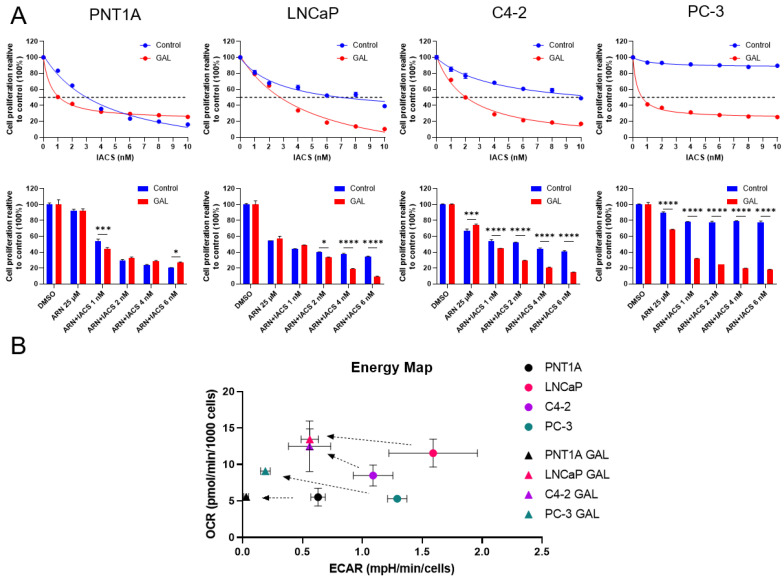
Prostate cancer (PCa) often becomes drug-treatment-resistant, posing a significant challenge to effective management. Although initial treatment with androgen deprivation therapy can control advanced PCa, subsequent resistance mechanisms allow tumor cells to continue growing, necessitating alternative approaches. This study delves into the specific metabolic dependencies of different PCa subtypes and explores the potential synergistic effects of combining androgen receptor (AR) inhibition (ARN with mitochondrial complex I inhibition (IACS)). We examined the metabolic behaviors of normal prostate epithelial cells (PNT1A), androgen-sensitive cells (LNCaP and C4-2), and androgen-independent cells (PC-3) when treated with ARN, IACS, or a combination. The results uncovered distinct mitochondrial activities across PCa subtypes, with androgen-dependent cells exhibiting heightened oxidative phosphorylation (OXPHOS). The combination of ARN and IACS significantly curbed cell proliferation in multiple PCa cell lines. Cellular bioenergetics analysis revealed that IACS reduced OXPHOS, while ARN hindered glycolysis in certain PCa cells. Additionally, galactose supplementation disrupted compensatory glycolytic mechanisms induced by metabolic reprogramming. Notably, glucose-deprived conditions heightened the sensitivity of PCa cells to mitochondrial inhibition, especially in the resistant PC-3 cells. Overall, this study illuminates the intricate interplay between AR signaling, metabolic adaptations, and treatment resistance in PCa. The findings offer valuable insights into subtype-specific metabolic profiles and propose a promising strategy to target PCa cells by exploiting their metabolic vulnerabilities.
Keywords: IACS-010759; apalutamide; metabolism; mitochondria; prostate cancer.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
Similar articles
-
Mitochondrial Elongation and ROS-Mediated Apoptosis in Prostate Cancer Cells under Therapy with Apalutamide and Complex I Inhibitor.Int J Mol Sci. 2024 Jun 25;25(13):6939. doi: 10.3390/ijms25136939. Int J Mol Sci. 2024. PMID: 39000047 Free PMC article.
-
Subtype and Site Specific-Induced Metabolic Vulnerabilities in Prostate Cancer.Mol Cancer Res. 2023 Jan 3;21(1):51-61. doi: 10.1158/1541-7786.MCR-22-0250. Mol Cancer Res. 2023. PMID: 36112348 Free PMC article.
-
Androgen-induced expression of DRP1 regulates mitochondrial metabolic reprogramming in prostate cancer.Cancer Lett. 2020 Feb 28;471:72-87. doi: 10.1016/j.canlet.2019.12.017. Epub 2019 Dec 12. Cancer Lett. 2020. PMID: 31838085
-
Understanding the mechanisms of androgen deprivation resistance in prostate cancer at the molecular level.Eur Urol. 2015 Mar;67(3):470-9. doi: 10.1016/j.eururo.2014.09.049. Epub 2014 Oct 8. Eur Urol. 2015. PMID: 25306226 Free PMC article. Review.
-
Androgen receptor (AR) heterogeneity in prostate cancer and therapy resistance.Cancer Lett. 2021 Oct 10;518:1-9. doi: 10.1016/j.canlet.2021.06.006. Epub 2021 Jun 10. Cancer Lett. 2021. PMID: 34118355 Free PMC article. Review.
Cited by
-
Mitochondrial Elongation and ROS-Mediated Apoptosis in Prostate Cancer Cells under Therapy with Apalutamide and Complex I Inhibitor.Int J Mol Sci. 2024 Jun 25;25(13):6939. doi: 10.3390/ijms25136939. Int J Mol Sci. 2024. PMID: 39000047 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
