

SMALL PLANT AND ORGAN 1 (SPO1) Encoding a Cellulose Synthase-like Protein D4 (OsCSLD4) Is an Important Regulator for Plant Architecture and Organ Size in Rice
- PMID: 38069299
- PMCID: PMC10707047
- DOI: 10.3390/ijms242316974
SMALL PLANT AND ORGAN 1 (SPO1) Encoding a Cellulose Synthase-like Protein D4 (OsCSLD4) Is an Important Regulator for Plant Architecture and Organ Size in Rice
Abstract
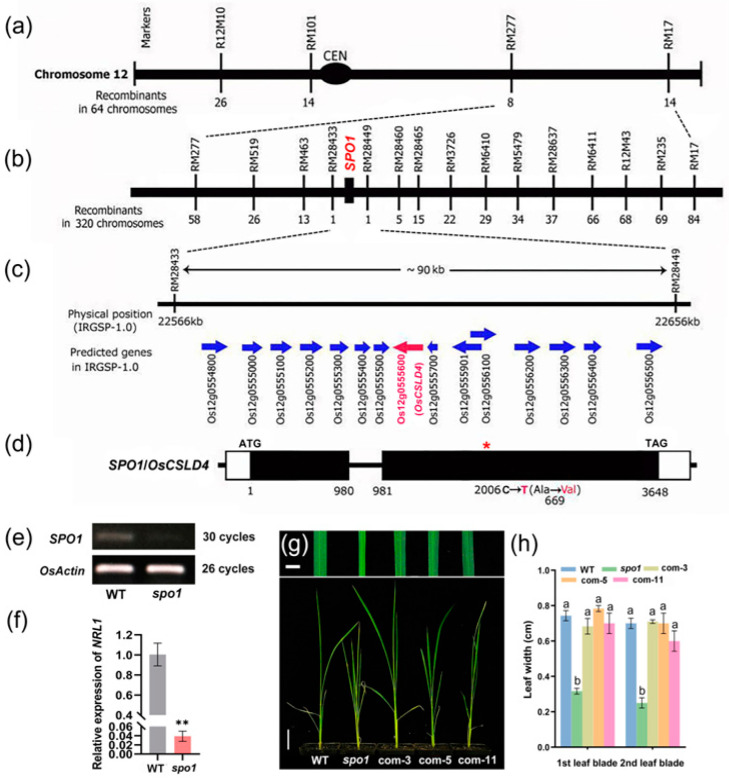
Plant architecture and organ size are considered as important traits in crop breeding and germplasm improvement. Although several factors affecting plant architecture and organ size have been identified in rice, the genetic and regulatory mechanisms remain to be elucidated. Here, we identified and characterized the small plant and organ 1 (spo1) mutant in rice (Oryza sativa), which exhibits narrow and rolled leaf, reductions in plant height, root length, and grain width, and other morphological defects. Map-based cloning revealed that SPO1 is allelic with OsCSLD4, a gene encoding the cellulose synthase-like protein D4, and is highly expressed in the roots at the seedling and tillering stages. Microscopic observation revealed the spo1 mutant had reduced number and width in leaf veins, smaller size of leaf bulliform cells, reduced cell length and cell area in the culm, and decreased width of epidermal cells in the outer glume of the grain. These results indicate the role of SPO1 in modulating cell division and cell expansion, which modulates plant architecture and organ size. It is showed that the contents of endogenous hormones including auxin, abscisic acid, gibberellin, and zeatin tested in the spo1 mutant were significantly altered, compared to the wild type. Furthermore, the transcriptome analysis revealed that the differentially expressed genes (DEGs) are significantly enriched in the pathways associated with plant hormone signal transduction, cell cycle progression, and cell wall formation. These results indicated that the loss of SPO1/OsCSLD4 function disrupted cell wall cellulose synthase and hormones homeostasis and signaling, thus leading to smaller plant and organ size in spo1. Taken together, we suggest the functional role of SPO1/OsCSLD4 in the control of rice plant and organ size by modulating cell division and expansion, likely through the effects of multiple hormonal pathways on cell wall formation.
Keywords: SPO1/OsCSLD4; cell division and expansion; narrow and rolled leaf; organ size; plant architecture; plant hormone.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Breuninger H., Lenhard M. Plant Development. Elsevier; Amsterdam, The Netherlands: 2010. Control of Tissue and Organ Growth in Plants; pp. 185–220. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
