

Tau seeding without tauopathy
- PMID: 38072056
- PMCID: PMC10797195
- DOI: 10.1016/j.jbc.2023.105545
Tau seeding without tauopathy
Abstract
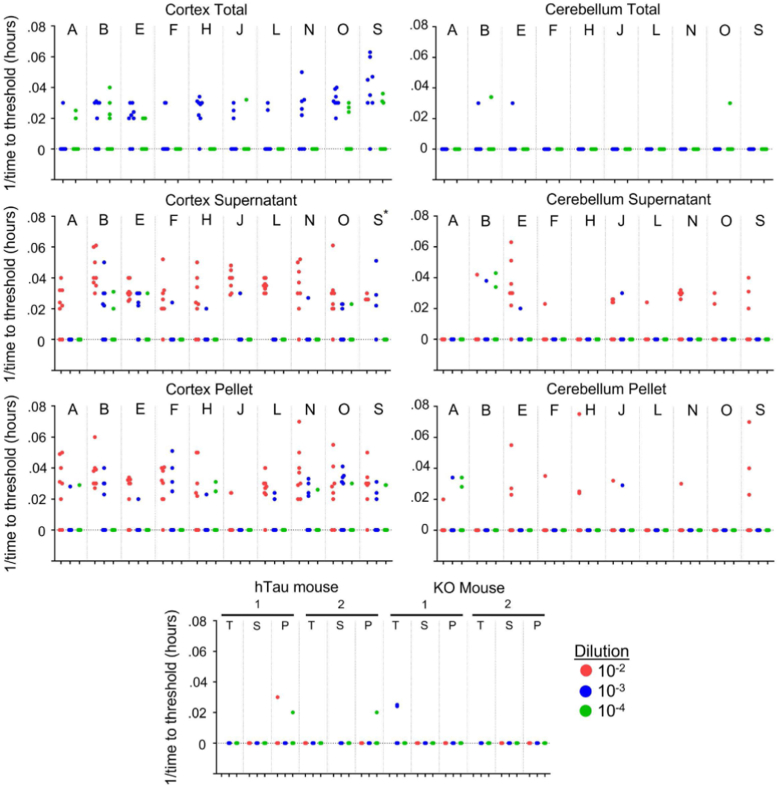
Neurodegenerative tauopathies such as Alzheimer's disease (AD) are caused by brain accumulation of tau assemblies. Evidence suggests tau functions as a prion, and cells and animals can efficiently propagate unique, transmissible tau pathologies. This suggests a dedicated cellular replication machinery, potentially reflecting a normal physiologic function for tau seeds. Consequently, we hypothesized that healthy control brains would contain seeding activity. We have recently developed a novel monoclonal antibody (MD3.1) specific for tau seeds. We used this antibody to immunopurify tau from the parietal and cerebellar cortices of 19 healthy subjects without any neuropathology, ranging 19 to 65 years. We detected seeding in lysates from the parietal cortex, but not in the cerebellum. We also detected no seeding in brain homogenates from wildtype or human tau knockin mice, suggesting that cellular/genetic context dictates development of seed-competent tau. Seeding did not correlate with subject age or brain tau levels. We confirmed our essential findings using an orthogonal assay, real-time quaking-induced conversion, which amplifies tau seeds in vitro. Dot blot analyses revealed no AT8 immunoreactivity above background levels in parietal and cerebellar extracts and ∼1/100 of that present in AD. Based on binding to a panel of antibodies, the conformational characteristics of control seeds differed from AD, suggesting a unique underlying assembly, or structural ensemble. Tau's ability to adopt self-replicating conformations under nonpathogenic conditions may reflect a normal function that goes awry in disease states.
Keywords: Alzheimer’s disease; FRET biosensor; healthy brain; prion; tau seeding activity; tauopathy.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures








References
-
- Lee V.M., Trojanowski J.Q. Neurodegenerative tauopathies: human disease and transgenic mouse models. Neuron. 1999;24:507–510. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
