

Uropathogenic Escherichia coli wield enterobactin-derived catabolites as siderophores
- PMID: 38072063
- PMCID: PMC10788543
- DOI: 10.1016/j.jbc.2023.105554
Uropathogenic Escherichia coli wield enterobactin-derived catabolites as siderophores
Abstract
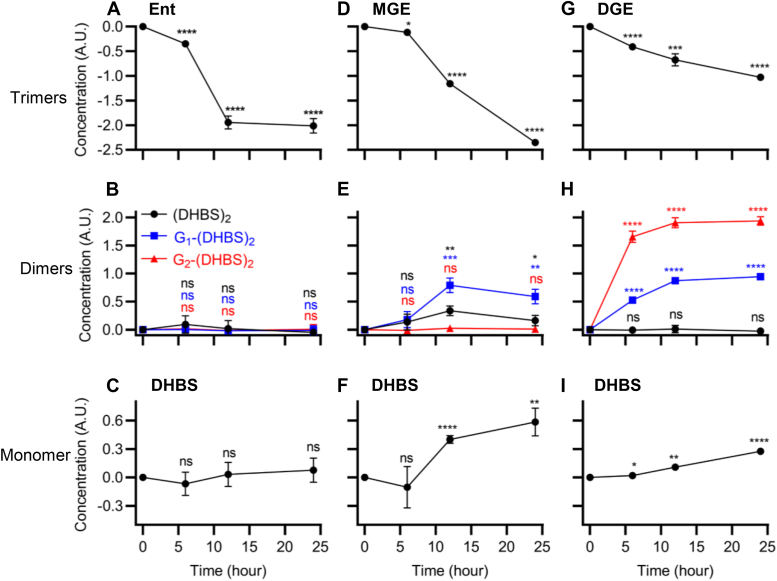
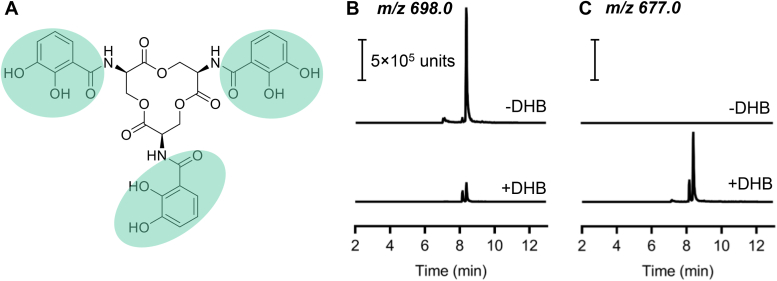
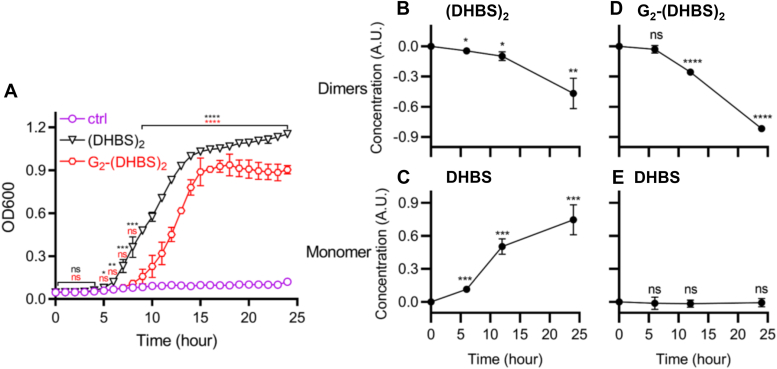
Uropathogenic Escherichia coli (UPEC) secrete multiple siderophore types to scavenge extracellular iron(III) ions during clinical urinary tract infections, despite the metabolic costs of biosynthesis. Here, we find the siderophore enterobactin (Ent) and its related products to be prominent components of the iron-responsive extracellular metabolome of a model UPEC strain. Using defined Ent biosynthesis and import mutants, we identify lower molecular weight dimeric exometabolites as products of incomplete siderophore catabolism, rather than prematurely released biosynthetic intermediates. In E. coli, iron acquisition from iron(III)-Ent complexes requires intracellular esterases that hydrolyze the siderophore. Although UPEC are equipped to consume the products of completely hydrolyzed Ent, we find that Ent and its derivatives may be incompletely hydrolyzed to yield products with retained siderophore activity. These results are consistent with catabolic inefficiency as means to obtain more than one iron ion per siderophore molecule. This is compatible with an evolved UPEC strategy to maximize the nutritional returns from metabolic investments in siderophore biosynthesis.
Keywords: Escherichia coli; Gram-negative bacteria; enterobactin; exometabolome; iron; metabolomics; siderophore.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
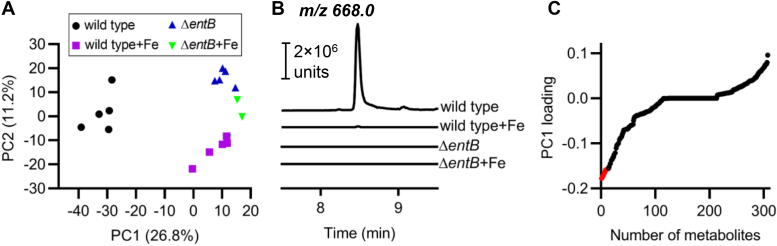
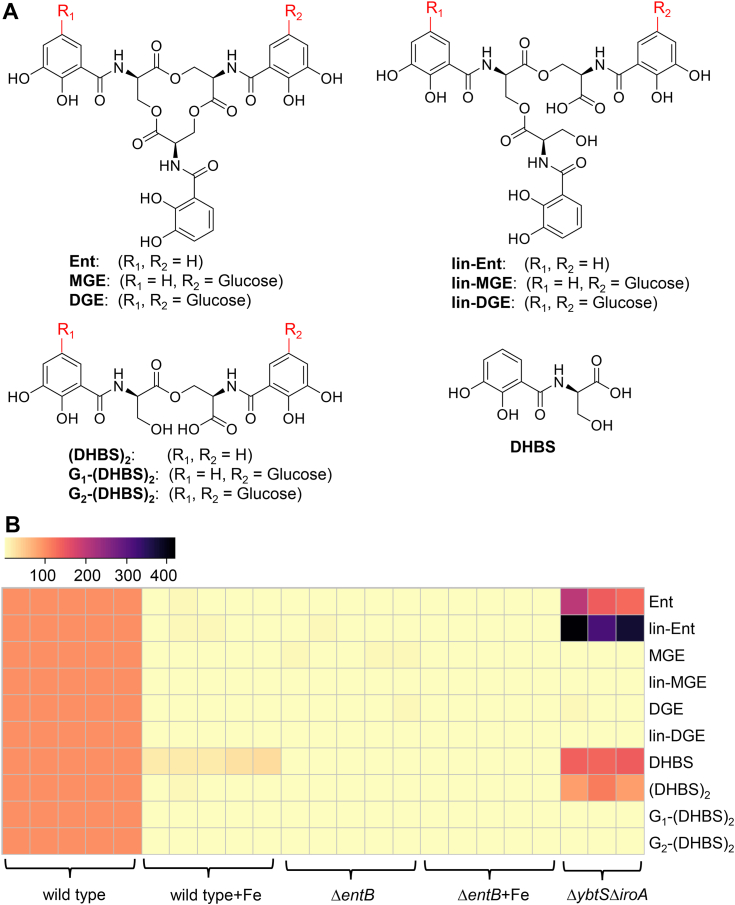
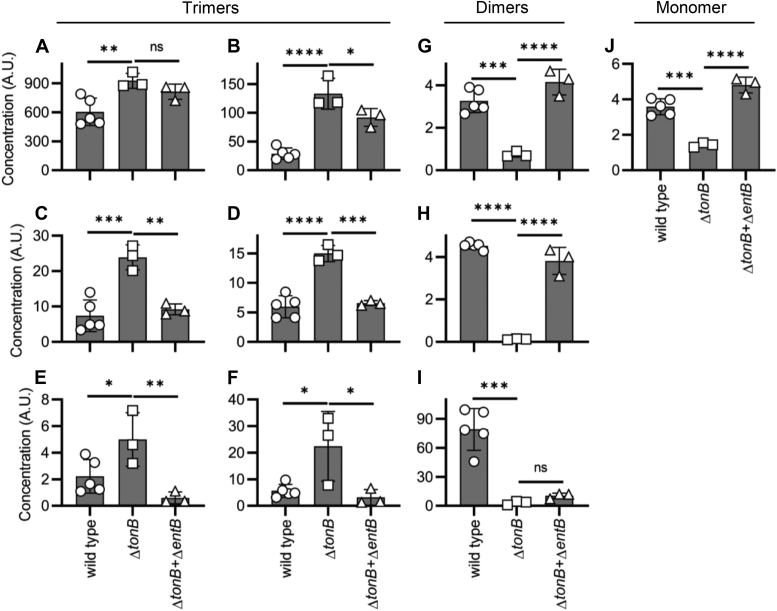
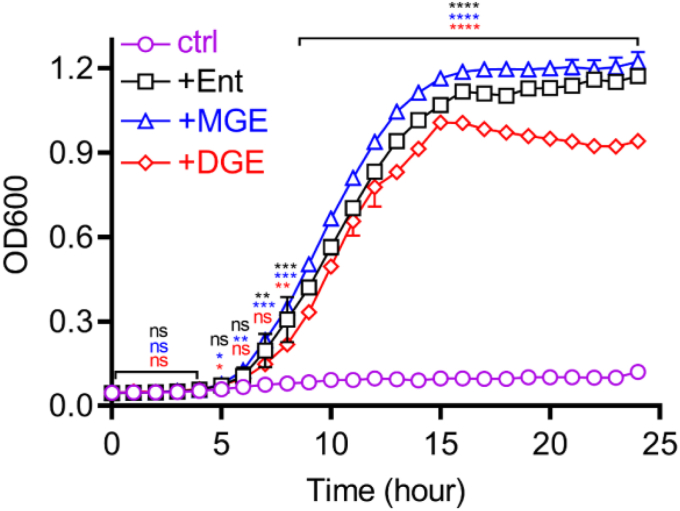
Update of
-
Uropathogenic Escherichia coli wield enterobactin-derived catabolites as siderophores.bioRxiv [Preprint]. 2023 Jul 25:2023.07.25.550588. doi: 10.1101/2023.07.25.550588. bioRxiv. 2023. Update in: J Biol Chem. 2024 Jan;300(1):105554. doi: 10.1016/j.jbc.2023.105554. PMID: 37546885 Free PMC article. Updated. Preprint.
References
-
- Trautner B.W. Urinary tract infections as a continuum: implications for diagnostic and antibiotic stewardship. Clin. Infect. Dis. 2021;72:1339–1341. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
