

This is a preprint.
CTCF-mediated 3D chromatin predetermines the gene expression program in the male germline
- PMID: 38076840
- PMCID: PMC10705413
- DOI: 10.1101/2023.11.30.569508
CTCF-mediated 3D chromatin predetermines the gene expression program in the male germline
Update in
-
CTCF-mediated 3D chromatin sets up the gene expression program in the male germline.Nat Struct Mol Biol. 2025 Jul;32(7):1227-1240. doi: 10.1038/s41594-025-01482-z. Epub 2025 Mar 3. Nat Struct Mol Biol. 2025. PMID: 40033153 Free PMC article.
Abstract
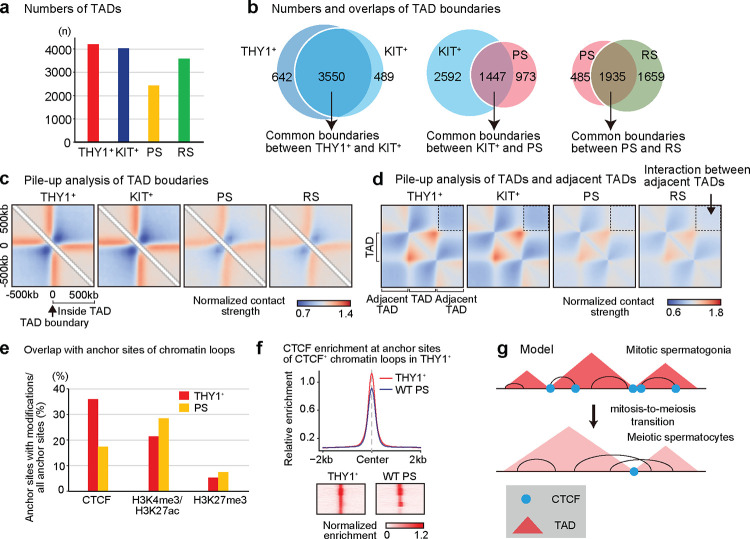
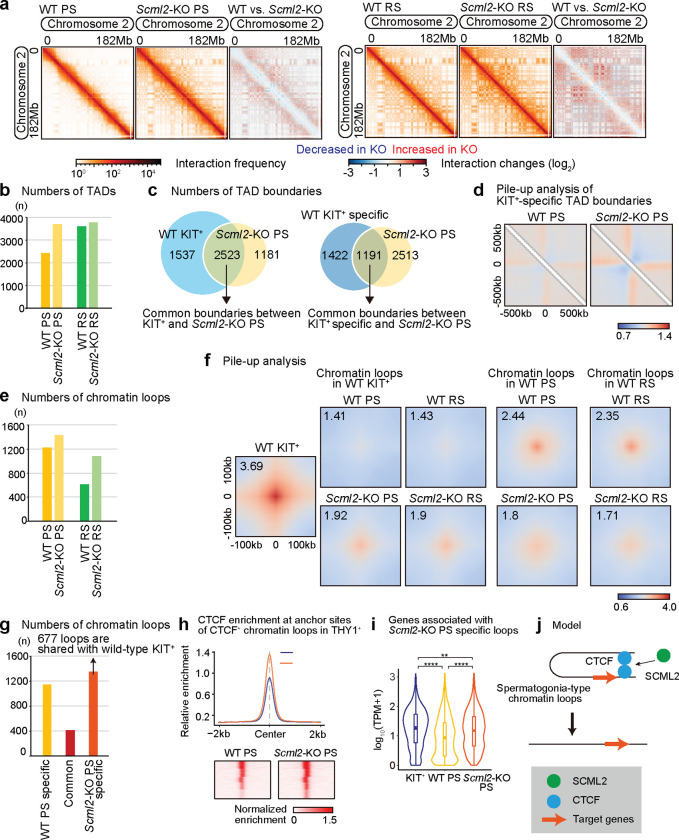
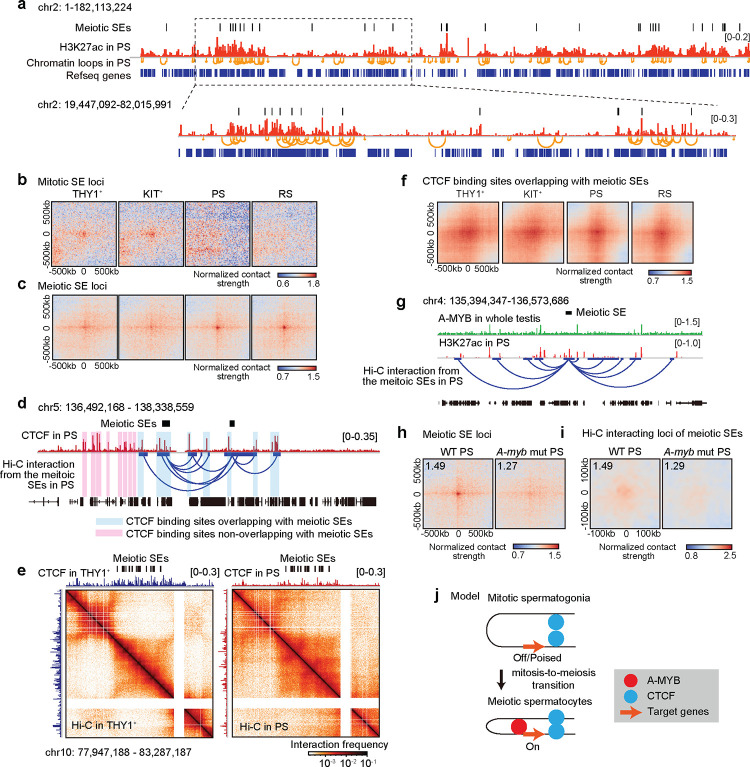
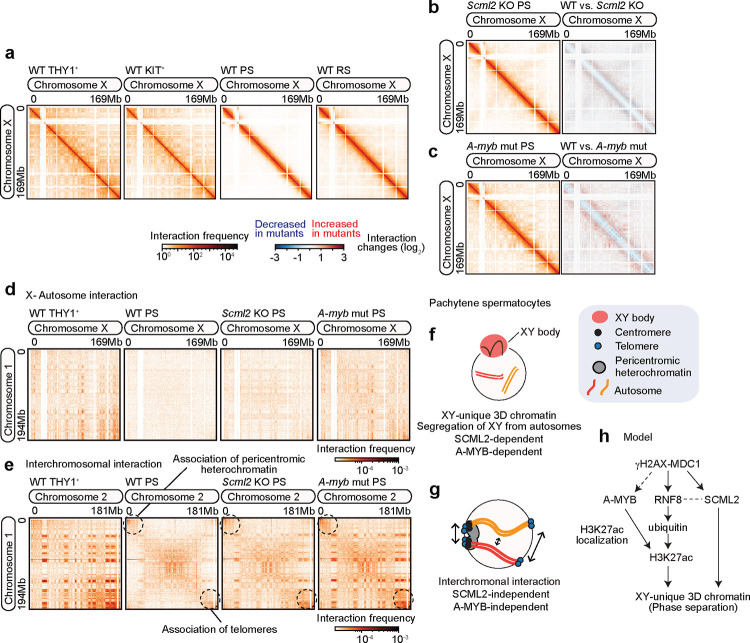
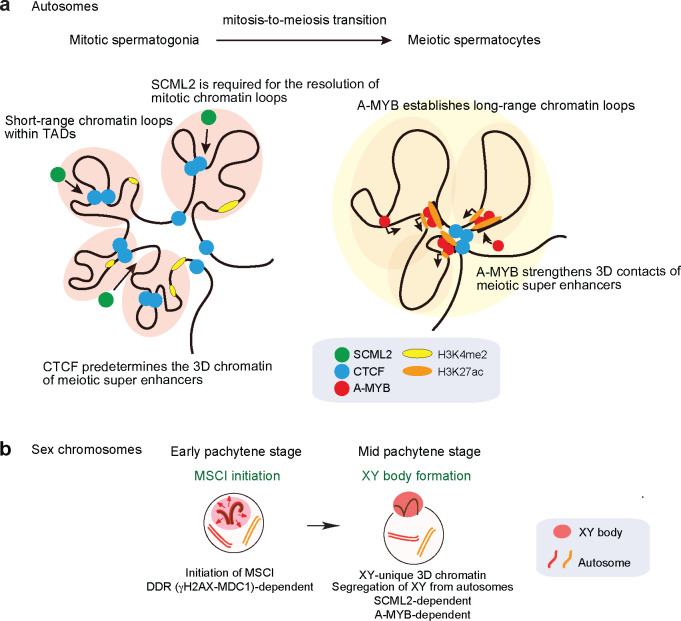
Spermatogenesis is a unidirectional differentiation process that generates haploid sperm, but how the gene expression program that directs this process is established is largely unknown. Here we determine the high-resolution 3D chromatin architecture of male germ cells during spermatogenesis and show that CTCF-mediated 3D chromatin predetermines the gene expression program required for spermatogenesis. In undifferentiated spermatogonia, CTCF-mediated chromatin contacts on autosomes pre-establish meiosis-specific super-enhancers (SE). These meiotic SE recruit the master transcription factor A-MYB in meiotic spermatocytes, which strengthens their 3D contacts and instructs a burst of meiotic gene expression. We also find that at the mitosis-to-meiosis transition, the germline-specific Polycomb protein SCML2 resolves chromatin loops that are specific to mitotic spermatogonia. Moreover, SCML2 and A-MYB establish the unique 3D chromatin organization of sex chromosomes during meiotic sex chromosome inactivation. We propose that CTCF-mediated 3D chromatin organization enforces epigenetic priming that directs unidirectional differentiation, thereby determining the cellular identity of the male germline.
Conflict of interest statement
Competing interest statement The authors declare no competing interests.
Figures


References
-
- Zheng H. & Xie W. The role of 3D genome organization in development and cell differentiation. Nat Rev Mol Cell Biol 20, 535–550 (2019). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases