

Gankyrin inhibits ferroptosis through the p53/SLC7A11/GPX4 axis in triple-negative breast cancer cells
- PMID: 38081931
- PMCID: PMC10713534
- DOI: 10.1038/s41598-023-49136-8
Gankyrin inhibits ferroptosis through the p53/SLC7A11/GPX4 axis in triple-negative breast cancer cells
Abstract
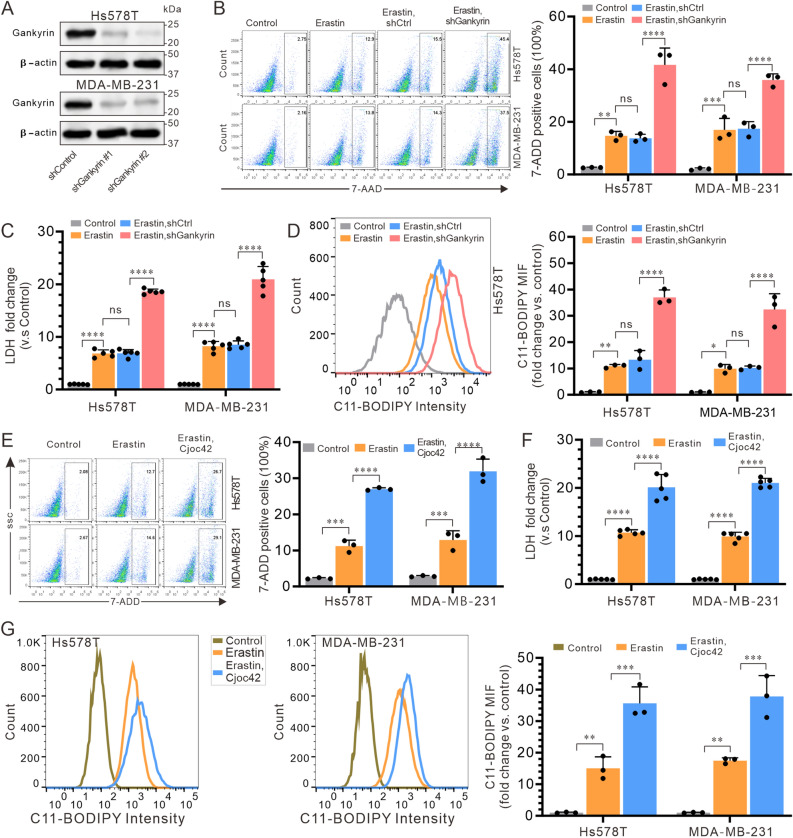
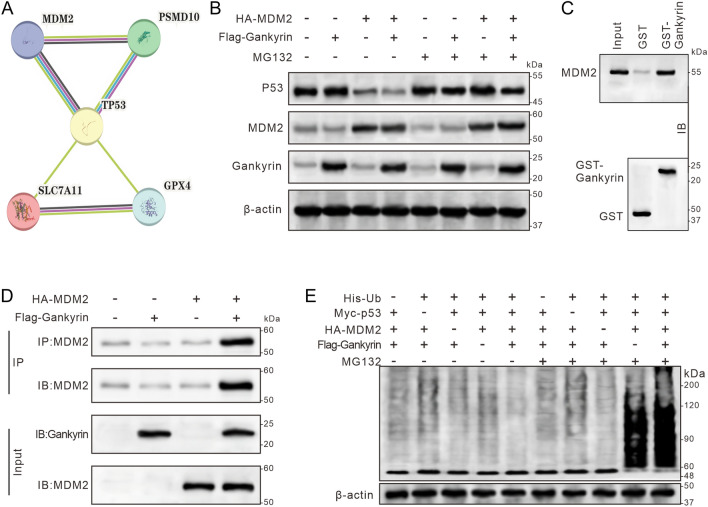
Gankyrin is found in high levels in triple-negative breast cancer (TNBC) and has been established to form a complex with the E3 ubiquitin ligase MDM2 and p53, resulting in the degradation of p53 in hepatocarcinoma cells. Therefore, this study sought to determine whether gankyrin could inhibit ferroptosis through this mechanism in TNBC cells. The expression of gankyrin was investigated in relation to the prognosis of TNBC using bioinformatics. Co-immunoprecipitation and GST pull-down assays were then conducted to determine the presence of a gankyrin and MDM2 complex. RT-qPCR and immunoblotting were used to examine molecules related to ferroptosis, such as gankyrin, p53, MDM2, SLC7A11, and GPX4. Additionally, cell death was evaluated using flow cytometry detection of 7-AAD and a lactate dehydrogenase release assay, as well as lipid peroxide C11-BODIPY. Results showed that the expression of gankyrin is significantly higher in TNBC tissues and cell lines, and is associated with a poor prognosis for patients. Subsequent studies revealed that inhibiting gankyrin activity triggered ferroptosis in TNBC cells. Additionally, silencing gankyrin caused an increase in the expression of the p53 protein, without altering its mRNA expression. Co-immunoprecipitation and GST pull-down experiments indicated that gankyrin and MDM2 form a complex. In mouse embryonic fibroblasts lacking both MDM2 and p53, this gankyrin/MDM2 complex was observed to ubiquitinate p53, thus raising the expression of molecules inhibited by ferroptosis, such as SLC7A11 and GPX4. Furthermore, silencing gankyrin in TNBC cells disrupted the formation of the gankyrin/MDM2 complex, hindered the degradation of p53, increased SLC7A11 expression, impeded cysteine uptake, and decreased GPX4 production. Our findings suggest that TNBC cells are able to prevent cell ferroptosis through the gankyrin/p53/SLC7A11/GPX4 signaling pathway, indicating that gankyrin may be a useful biomarker for predicting TNBC prognosis or a potential therapeutic target.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
