

Age-Related Alterations in Peripheral Immune Landscape with Magnified Impact on Post-Stroke Brain
- PMID: 38090608
- PMCID: PMC10712880
- DOI: 10.34133/research.0287
Age-Related Alterations in Peripheral Immune Landscape with Magnified Impact on Post-Stroke Brain
Abstract
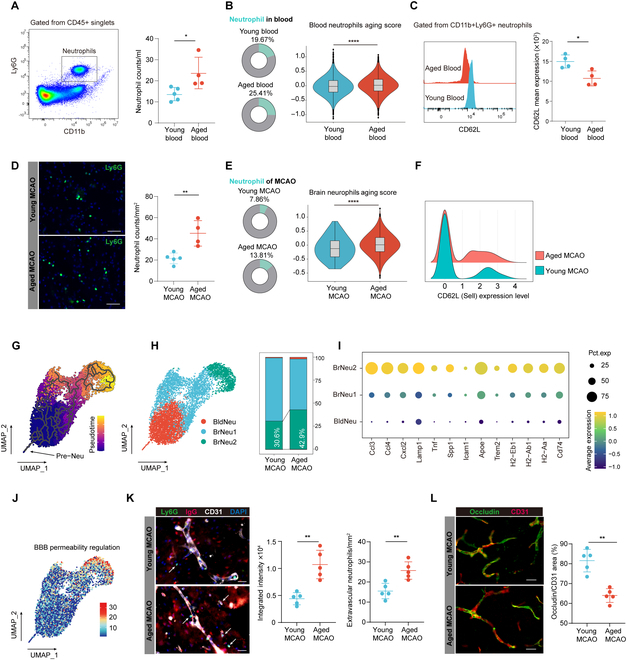
Immunosenescence refers to the multifaceted and profound alterations in the immune system brought about by aging, exerting complex influences on the pathophysiological processes of diseases that manifest upon it. Using a combination of single-cell RNA sequencing, cytometry by time of flight, and various immunological assays, we investigated the characteristics of immunosenescence in the peripheral blood of aged mice and its impact on the cerebral immune environment after ischemic stroke. Our results revealed some features of immunosenescence. We observed an increase in neutrophil counts, concurrent with accelerated neutrophil aging, characterized by altered expression of aging-associated markers like CD62L and consequential changes in neutrophil-mediated immune functions. Monocytes/macrophages in aged mice exhibited enhanced antigen-presentation capabilities. T cell profiles shifted from naive to effector or memory states, with a specific rise in T helper 1 cells and T helper 17 cells subpopulations and increased regulatory T cell activation in CD4 T cells. Furthermore, regulatory CD8 T cells marked by Klra decreased with aging, while a subpopulation of exhausted-like CD8 T cells expanded, retaining potent immunostimulatory and proinflammatory functions. Critically, these inherent disparities not only persisted but were further amplified within the ischemic hemispheres following stroke. In summary, our comprehensive insights into the key attributes of peripheral immunosenescence provide a vital theoretical foundation for understanding not only ischemic strokes but also other age-associated diseases.
Copyright © 2023 Jianan Lu et al.
Conflict of interest statement
Competing interests: The authors declare that they have no competing interests.
Figures





References
-
- Shichita T, Ooboshi H, Yoshimura A. Neuroimmune mechanisms and therapies mediating post-ischaemic brain injury and repair. Nat Rev Neurosci. 2023;24(5):299–312. - PubMed
-
- Prinz M, Priller J. The role of peripheral immune cells in the CNS in steady state and disease. Nat Neurosci. 2017;20(2):136–144. - PubMed
-
- Nikolich-Žugich J. The twilight of immunity: Emerging concepts in aging of the immune system. Nat Immunol. 2018;19(1):10–19. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
