

TCR signaling promotes formation of an STS1-Cbl-b complex with pH-sensitive phosphatase activity that suppresses T cell function in acidic environments
- PMID: 38091950
- PMCID: PMC10785950
- DOI: 10.1016/j.immuni.2023.11.010
TCR signaling promotes formation of an STS1-Cbl-b complex with pH-sensitive phosphatase activity that suppresses T cell function in acidic environments
Abstract
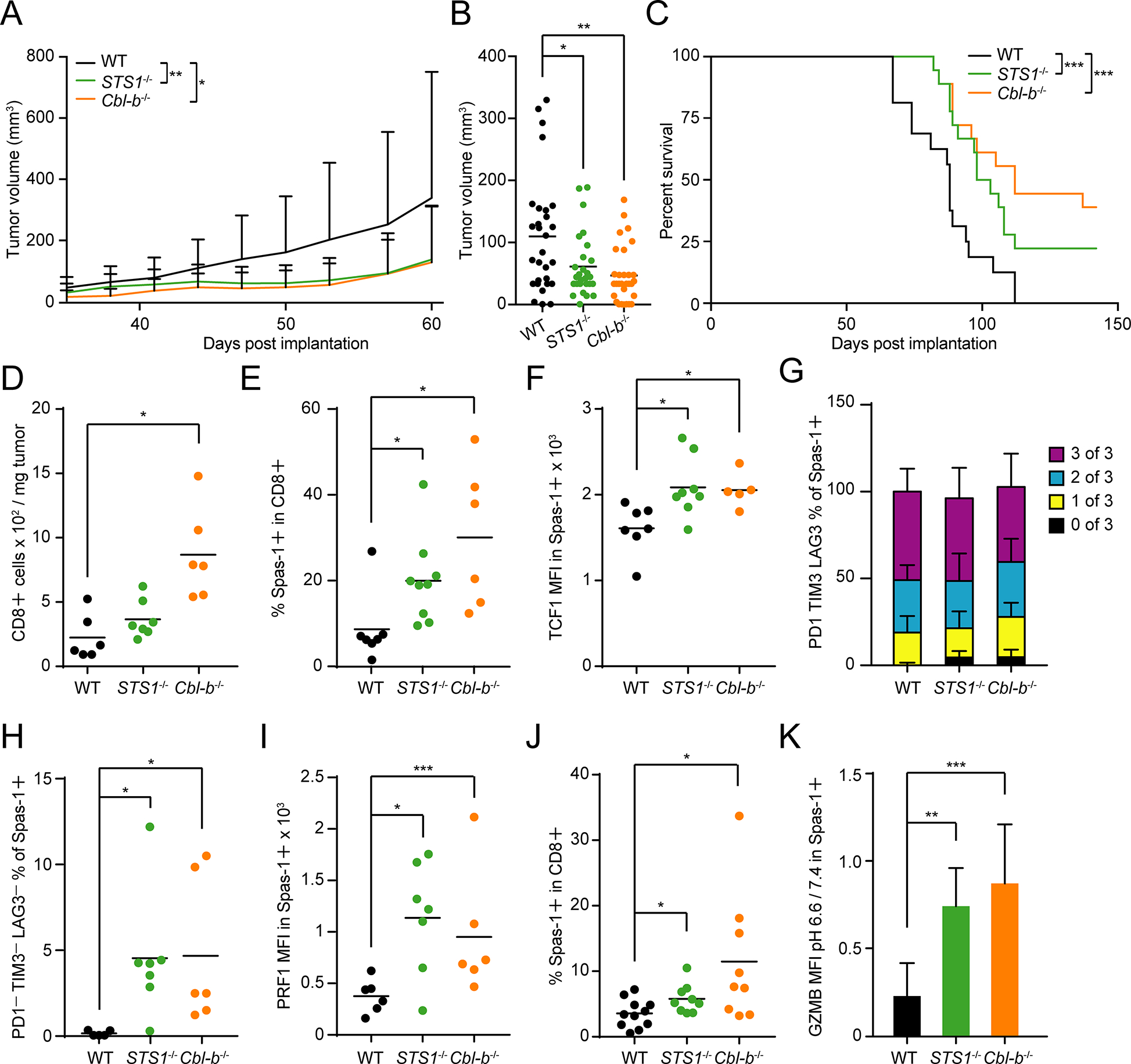
T cell responses are inhibited by acidic environments. T cell receptor (TCR)-induced protein phosphorylation is negatively regulated by dephosphorylation and/or ubiquitination, but the mechanisms underlying sensitivity to acidic environments are not fully understood. Here, we found that TCR stimulation induced a molecular complex of Cbl-b, an E3-ubiquitin ligase, with STS1, a pH-sensitive unconventional phosphatase. The induced interaction depended upon a proline motif in Cbl-b interacting with the STS1 SH3 domain. STS1 dephosphorylated Cbl-b interacting phosphoproteins. The deficiency of STS1 or Cbl-b diminished the sensitivity of T cell responses to the inhibitory effects of acid in an autocrine or paracrine manner in vitro or in vivo. Moreover, the deficiency of STS1 or Cbl-b promoted T cell proliferative and differentiation activities in vivo and inhibited tumor growth, prolonged survival, and improved T cell fitness in tumor models. Thus, a TCR-induced STS1-Cbl-b complex senses intra- or extra-cellular acidity and regulates T cell responses, presenting a potential therapeutic target for improving anti-tumor immunity.
Keywords: TCR signaling; pH sensing; phosphatase.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.W. is a co-founder and a scientific advisory board (SAB) member of Nurix Therapeutics, Inc., which has a Cbl-b inhibitor in phase I clinical trials. Dr. Weiss has founder shares and receives payment for his role on the Nurix SAB. A.W. also serves on the advisory boards of BlueSphere Bio, BridGene Biologics, EpiBiologics, Genentech, Imidomics, and Jasper Therapeutics. L.F. has received research support from Roche/Genentech, Abbvie, Bavarian Nordic, Bristol Myers Squibb, Dendreon, Janssen, Merck, and Partner Therapeutics. L.F. has served on the SABs of Actym, Astra Zeneca, Atreca, Bioatla, Bolt, Bristol Myer Squibb, Daiichi Sankyo, Immunogenesis, Innovent, Merck, Merck KGA, Nutcracker, RAPT, Scribe, Senti, Sutro, and Roche/Genentech.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
