

A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain
- PMID: 38092916
- PMCID: PMC10719114
- DOI: 10.1038/s41586-023-06812-z
A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain
Abstract
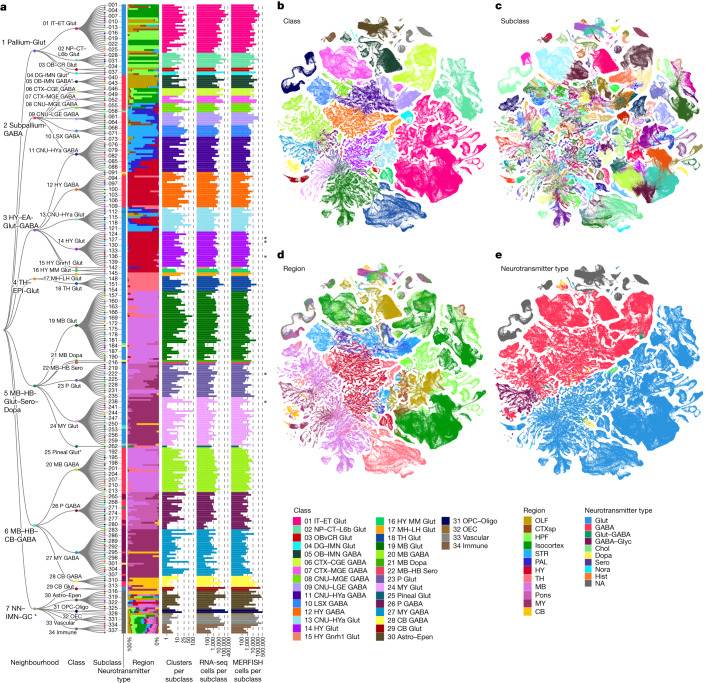
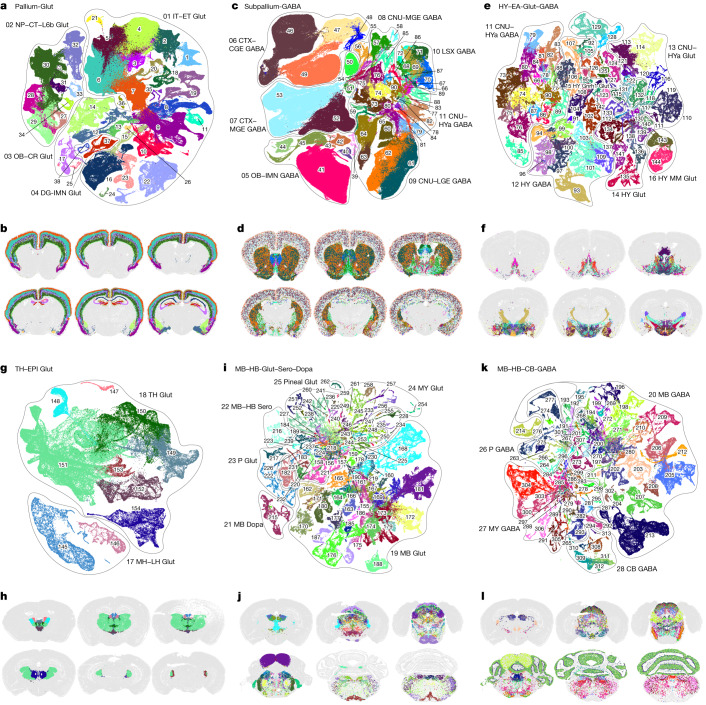
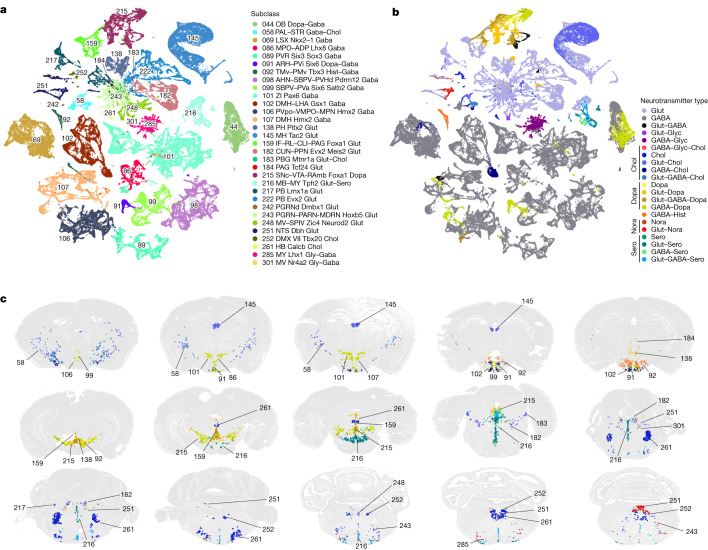
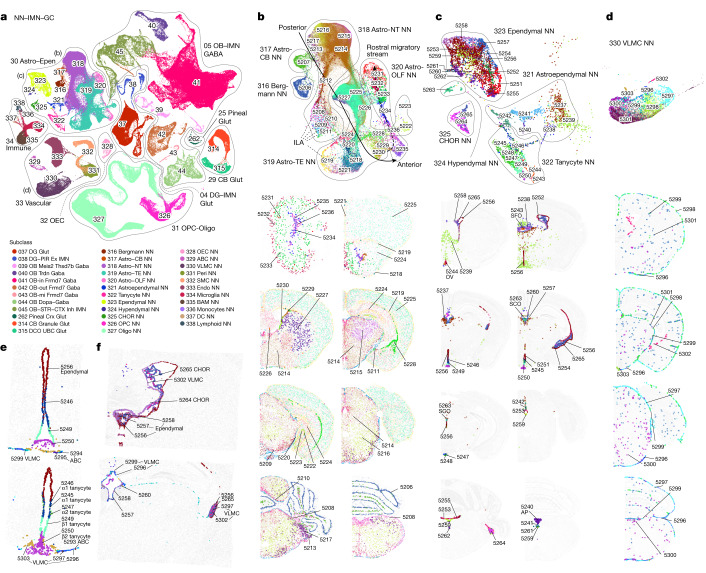
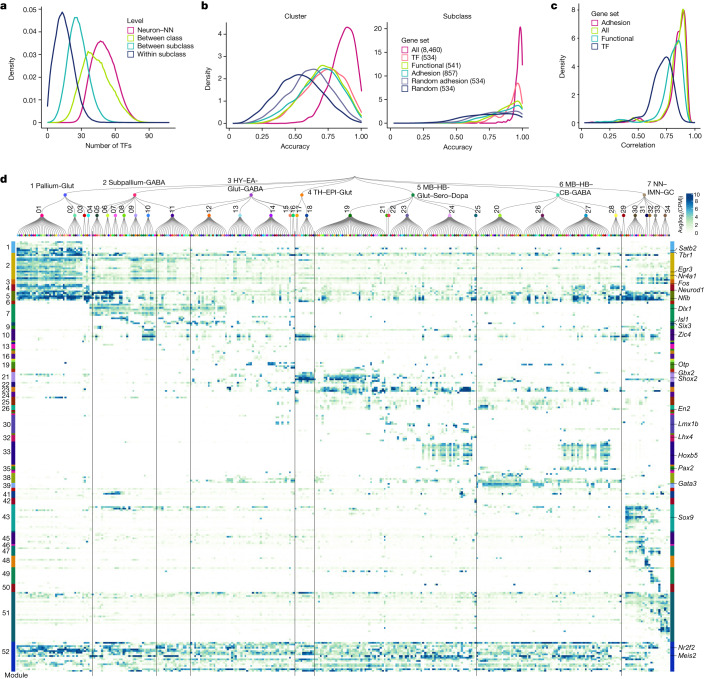
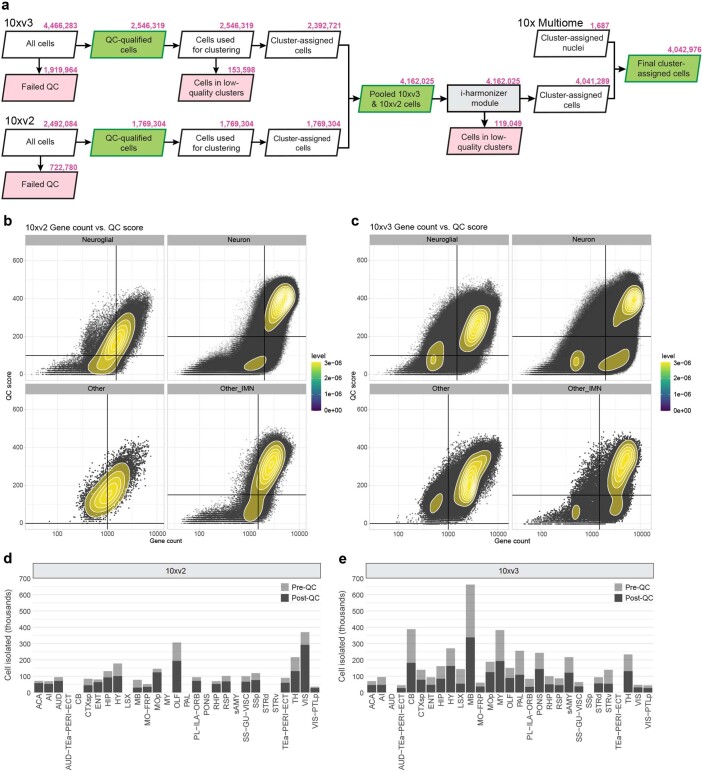
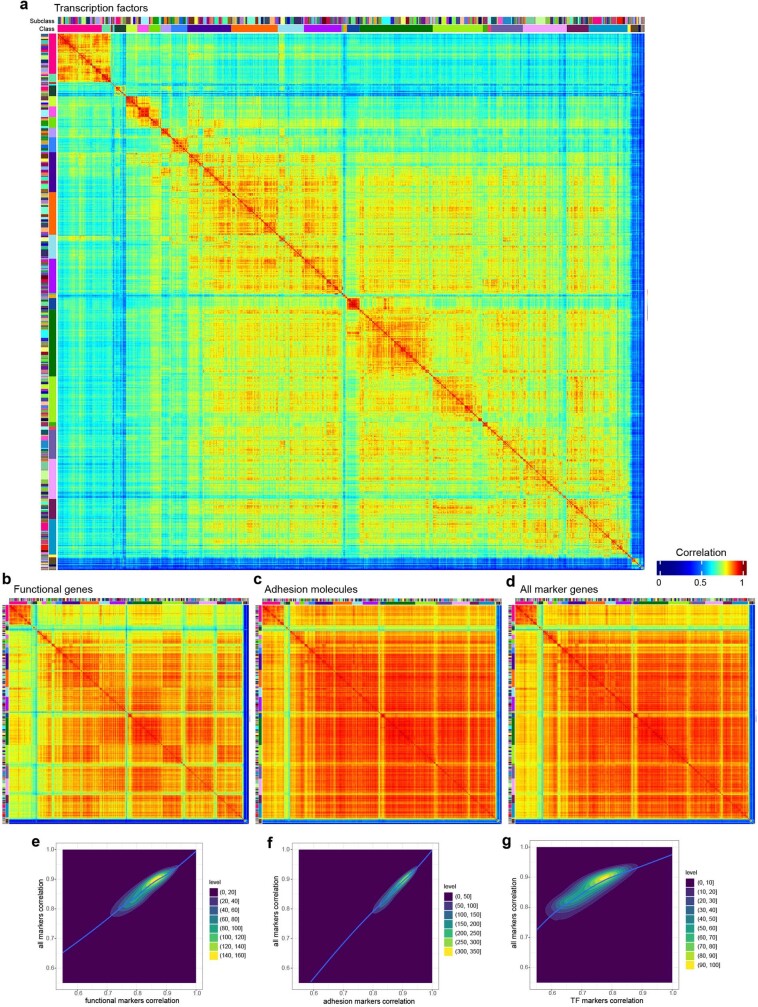
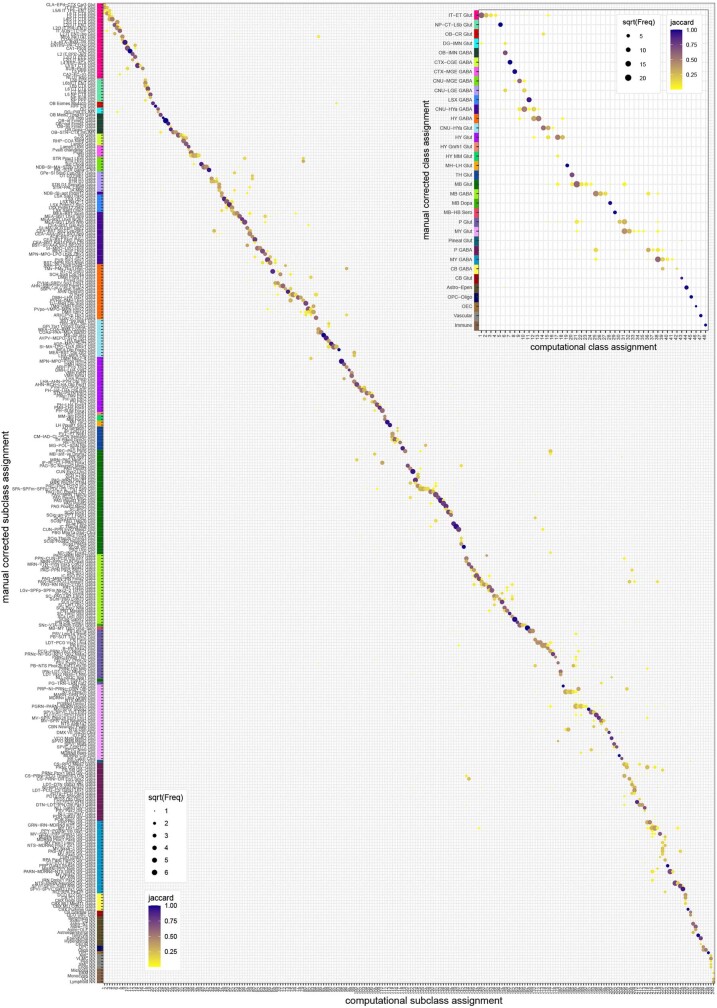
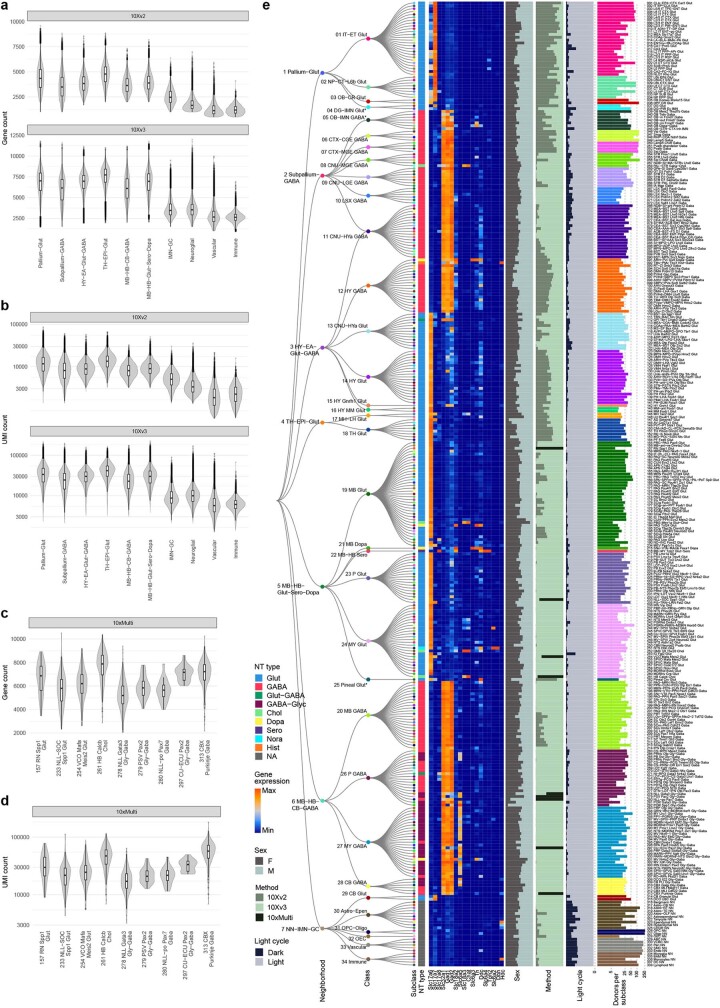
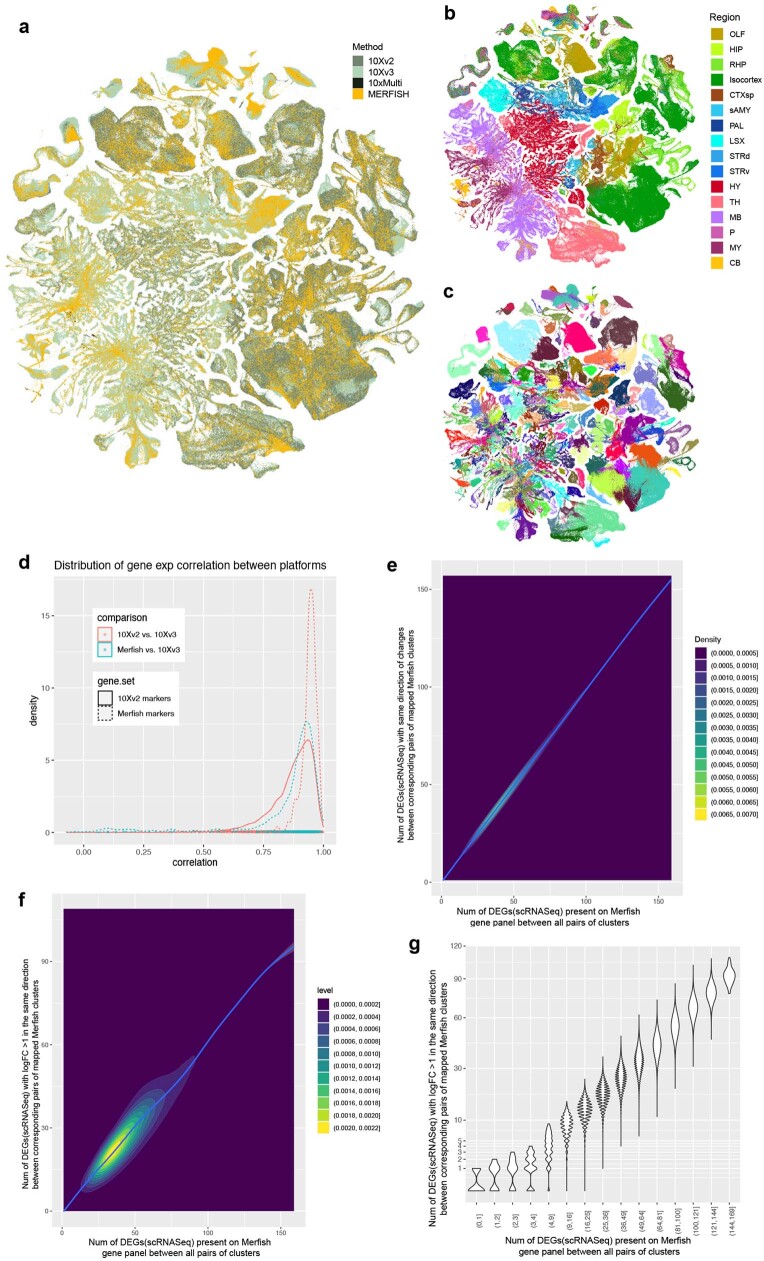
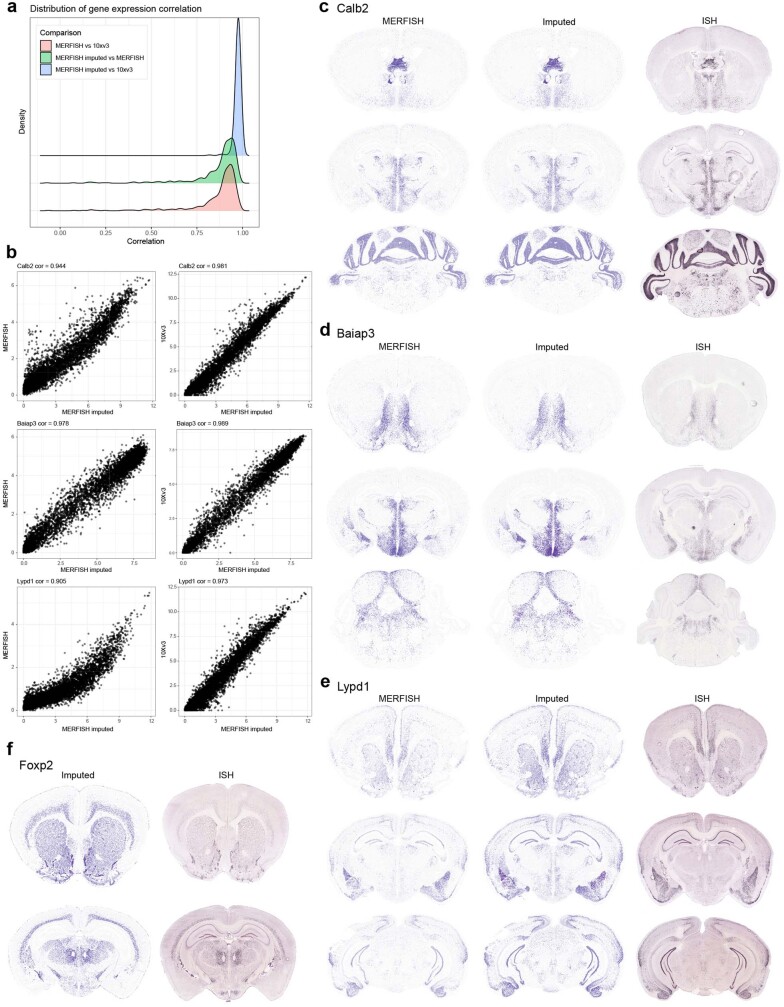
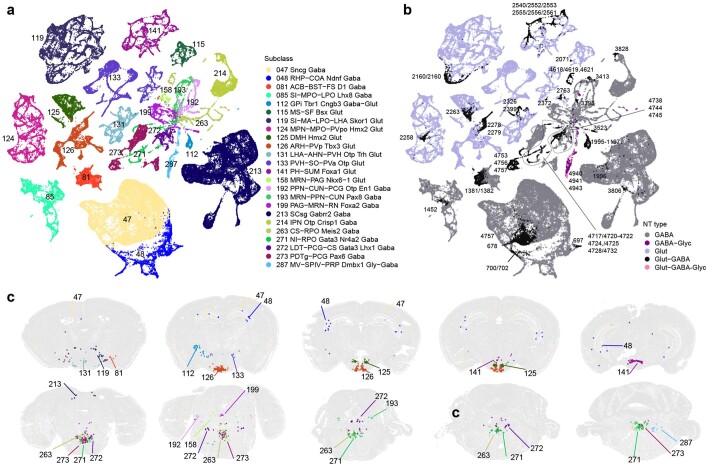
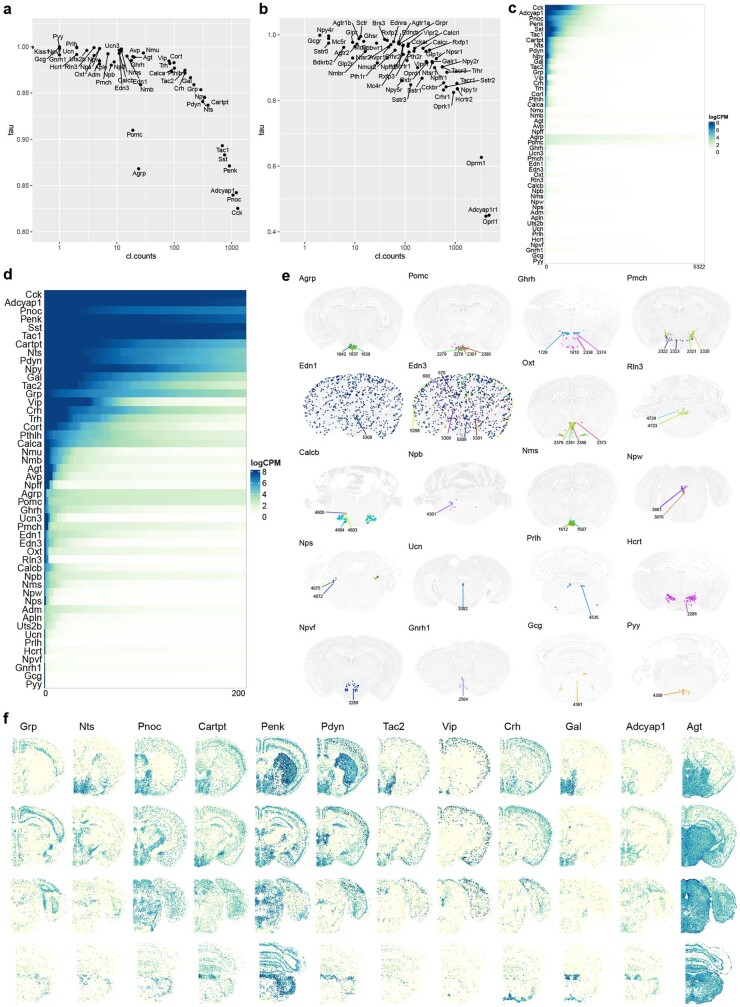
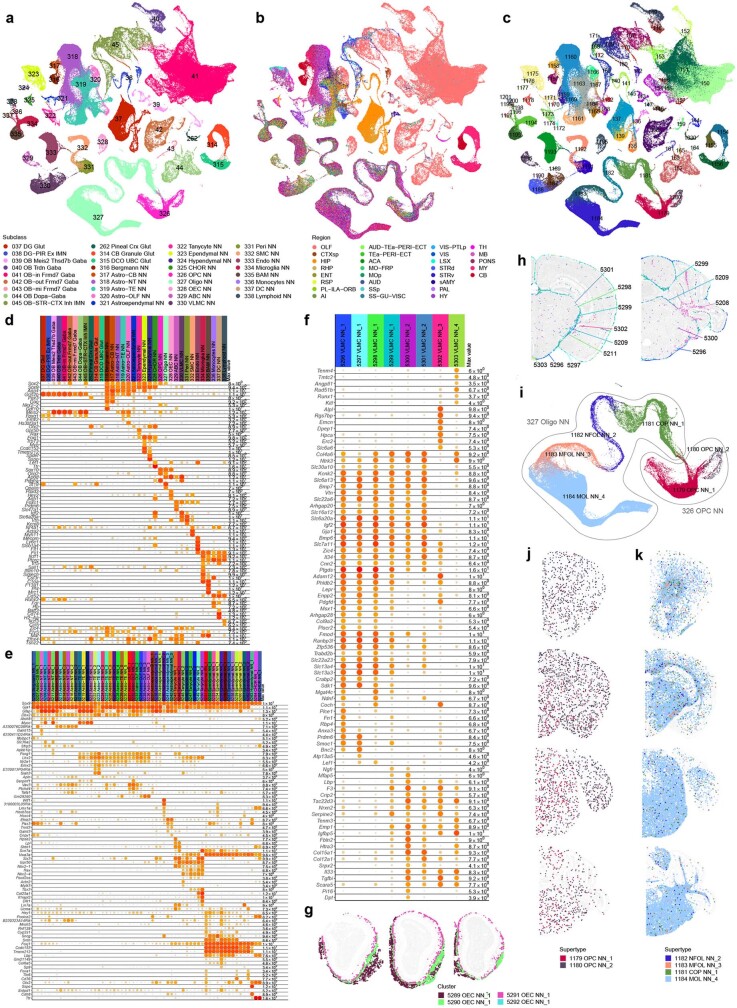
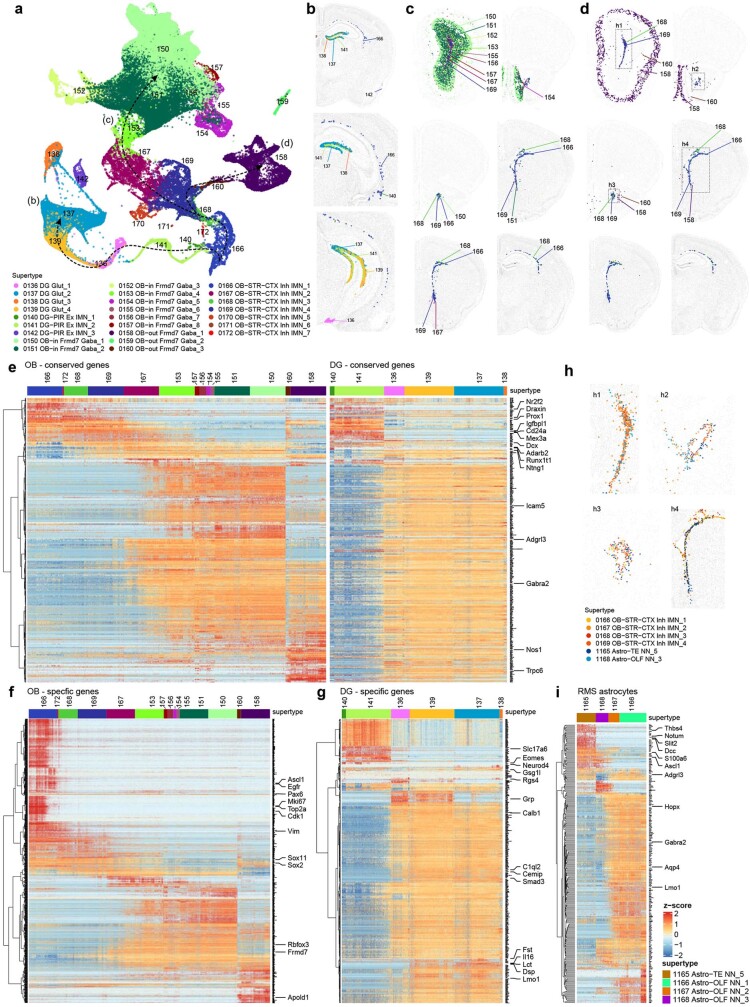
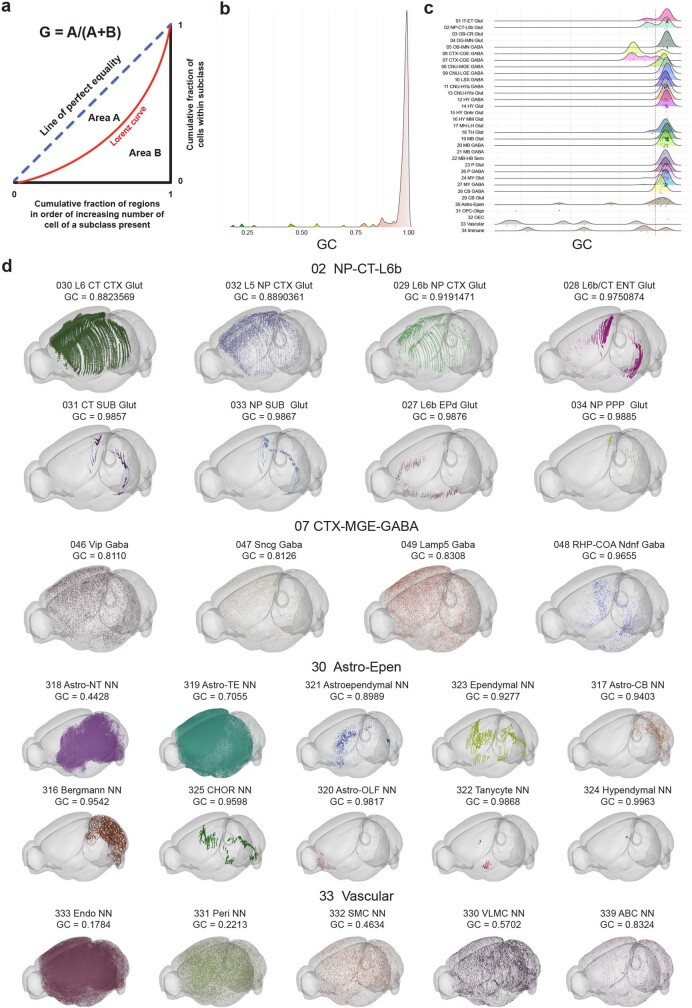
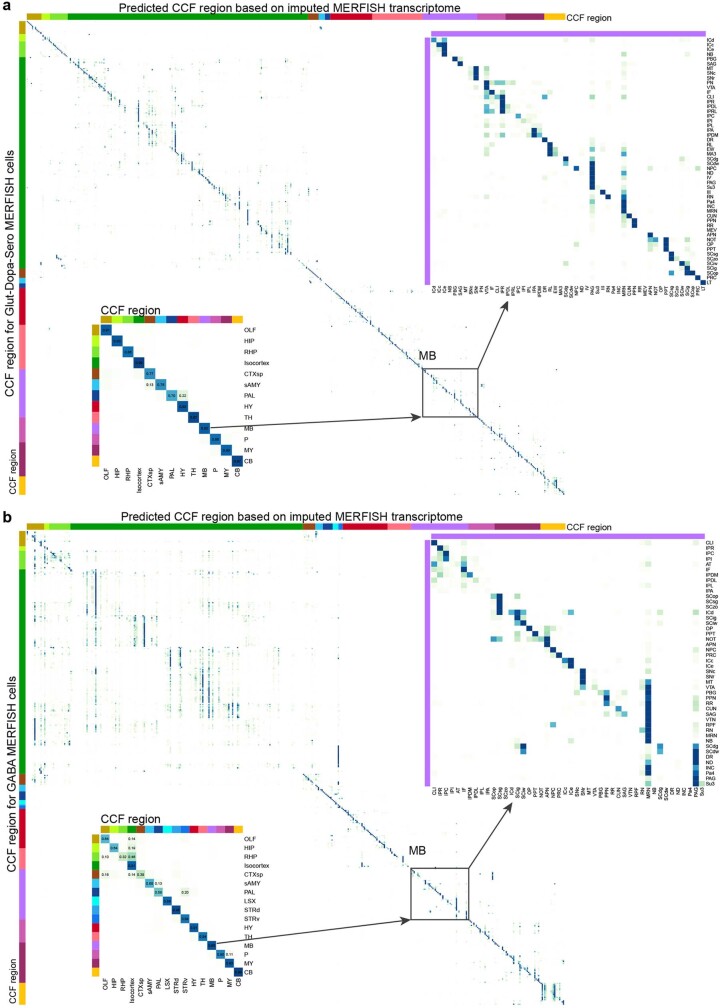
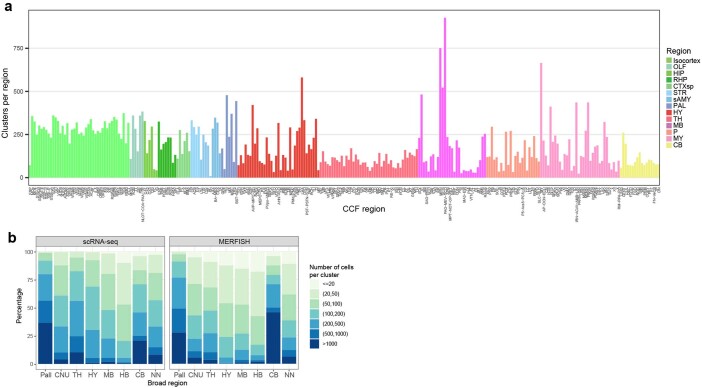
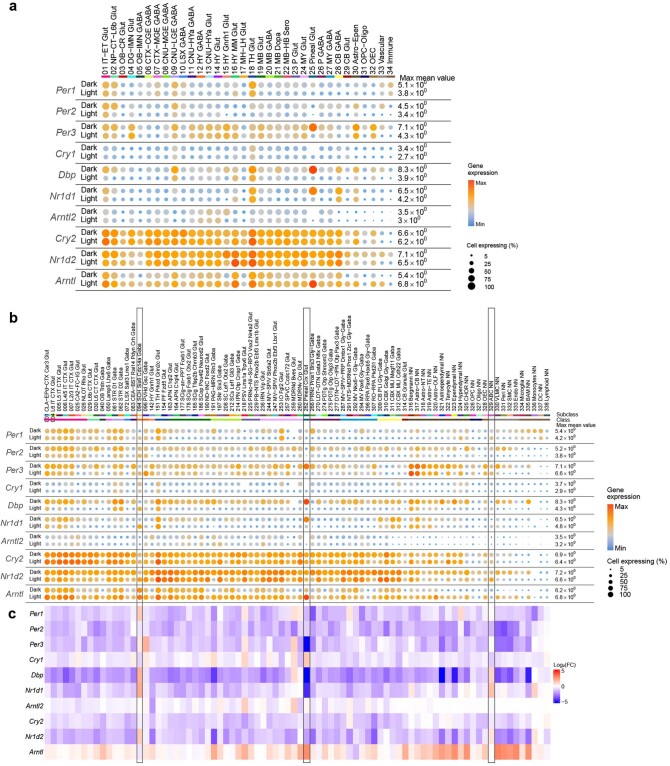
The mammalian brain consists of millions to billions of cells that are organized into many cell types with specific spatial distribution patterns and structural and functional properties1-3. Here we report a comprehensive and high-resolution transcriptomic and spatial cell-type atlas for the whole adult mouse brain. The cell-type atlas was created by combining a single-cell RNA-sequencing (scRNA-seq) dataset of around 7 million cells profiled (approximately 4.0 million cells passing quality control), and a spatial transcriptomic dataset of approximately 4.3 million cells using multiplexed error-robust fluorescence in situ hybridization (MERFISH). The atlas is hierarchically organized into 4 nested levels of classification: 34 classes, 338 subclasses, 1,201 supertypes and 5,322 clusters. We present an online platform, Allen Brain Cell Atlas, to visualize the mouse whole-brain cell-type atlas along with the single-cell RNA-sequencing and MERFISH datasets. We systematically analysed the neuronal and non-neuronal cell types across the brain and identified a high degree of correspondence between transcriptomic identity and spatial specificity for each cell type. The results reveal unique features of cell-type organization in different brain regions-in particular, a dichotomy between the dorsal and ventral parts of the brain. The dorsal part contains relatively fewer yet highly divergent neuronal types, whereas the ventral part contains more numerous neuronal types that are more closely related to each other. Our study also uncovered extraordinary diversity and heterogeneity in neurotransmitter and neuropeptide expression and co-expression patterns in different cell types. Finally, we found that transcription factors are major determinants of cell-type classification and identified a combinatorial transcription factor code that defines cell types across all parts of the brain. The whole mouse brain transcriptomic and spatial cell-type atlas establishes a benchmark reference atlas and a foundational resource for integrative investigations of cellular and circuit function, development and evolution of the mammalian brain.
© 2023. The Author(s).
Conflict of interest statement
H.Z. is on the scientific advisory board of MapLight Therapeutics, Inc. X.Z. is a co-founder of and consultant for Vizgen.
Figures





Update of
-
A high-resolution transcriptomic and spatial atlas of cell types in the whole mouse brain.bioRxiv [Preprint]. 2023 Mar 6:2023.03.06.531121. doi: 10.1101/2023.03.06.531121. bioRxiv. 2023. Update in: Nature. 2023 Dec;624(7991):317-332. doi: 10.1038/s41586-023-06812-z. PMID: 37034735 Free PMC article. Updated. Preprint.
References
-
- Paxinos, G. The Rat Nervous System 4th edn (Academic Press, 2014).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
