

Dynamic altruistic cooperation within breast tumors
- PMID: 38093346
- PMCID: PMC10720132
- DOI: 10.1186/s12943-023-01896-7
Dynamic altruistic cooperation within breast tumors
Abstract
Background: Social behaviors such as altruism, where one self-sacrifices for collective benefits, critically influence an organism's survival and responses to the environment. Such behaviors are widely exemplified in nature but have been underexplored in cancer cells which are conventionally seen as selfish competitive players. This multidisciplinary study explores altruism and its mechanism in breast cancer cells and its contribution to chemoresistance.
Methods: MicroRNA profiling was performed on circulating tumor cells collected from the blood of treated breast cancer patients. Cancer cell lines ectopically expressing candidate miRNA were used in co-culture experiments and treated with docetaxel. Ecological parameters like relative survival and relative fitness were assessed using flow cytometry. Functional studies and characterization performed in vitro and in vivo include proliferation, iTRAQ-mass spectrometry, RNA sequencing, inhibition by small molecules and antibodies, siRNA knockdown, CRISPR/dCas9 inhibition and fluorescence imaging of promoter reporter-expressing cells. Mathematical modeling based on evolutionary game theory was performed to simulate spatial organization of cancer cells.
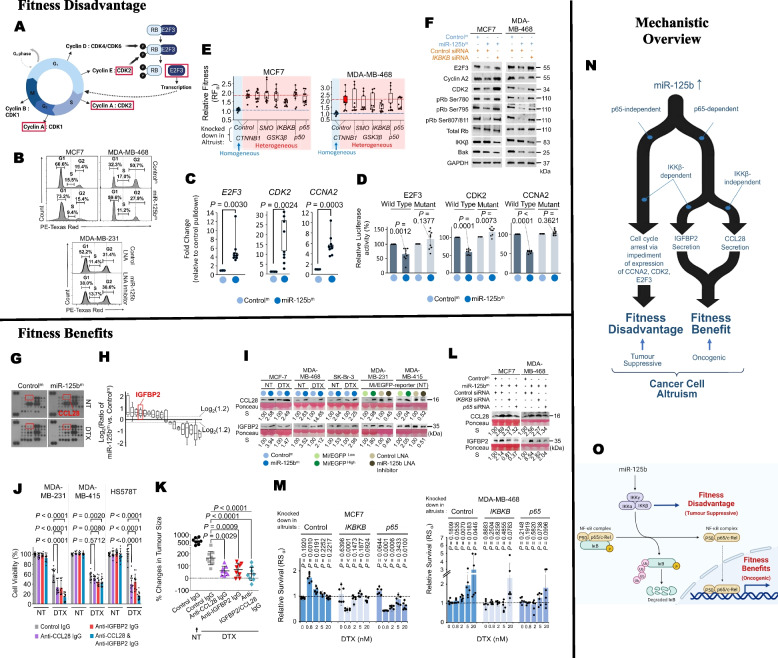
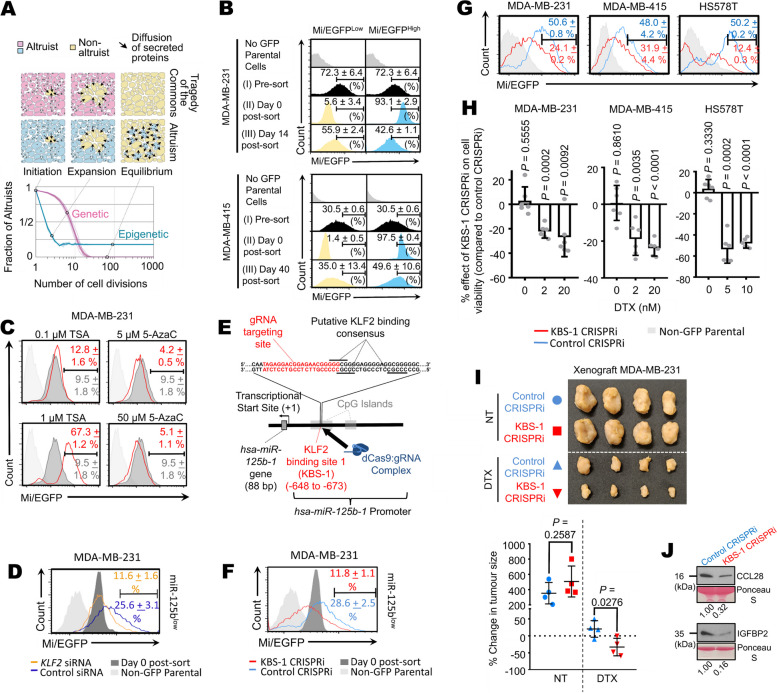
Results: Opposing cancer processes underlie altruism: an oncogenic process involving secretion of IGFBP2 and CCL28 by the altruists to induce survival benefits in neighboring cells under taxane exposure, and a self-sacrificial tumor suppressive process impeding proliferation of altruists via cell cycle arrest. Both processes are regulated concurrently in the altruists by miR-125b, via differential NF-κB signaling specifically through IKKβ. Altruistic cells persist in the tumor despite their self-sacrifice, as they can regenerate epigenetically from non-altruists via a KLF2/PCAF-mediated mechanism. The altruists maintain a sparse spatial organization by inhibiting surrounding cells from adopting the altruistic fate via a lateral inhibition mechanism involving a GAB1-PI3K-AKT-miR-125b signaling circuit.
Conclusions: Our data reveal molecular mechanisms underlying manifestation, persistence and spatial spread of cancer cell altruism. A minor population behave altruistically at a cost to itself producing a collective benefit for the tumor, suggesting tumors to be dynamic social systems governed by the same rules of cooperation in social organisms. Understanding cancer cell altruism may lead to more holistic models of tumor evolution and drug response, as well as therapeutic paradigms that account for social interactions. Cancer cells constitute tractable experimental models for fields beyond oncology, like evolutionary ecology and game theory.
© 2023. The Author(s).
Conflict of interest statement
Lihan Zhou and Mo-Huang Li are employed as Chief Executive Officers of MiRXES Pte Ltd and CellSievo Pte Ltd respectively.
Figures



References
-
- Miller BE, Miller FR, Wilburn D, Heppner GH. Dominance of a tumor subpopulation line in mixed heterogeneous mouse mammary tumors. Cancer Res. 1988;48:5747–5753. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- NUS Postgraduate Research Scholarships/National University of Singapore
- NUS Postgraduate Research Scholarships/National University of Singapore
- Department of Pathology Startup Grants/National University of Singapore
- Department of Pathology Startup Grants/National University of Singapore
- T1- NUHS Joint Grant Call FY17 - 2nd call-10/Ministry of Education - Singapore
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
