

Structural characterization of SLYM-a 4th meningeal membrane
- PMID: 38098084
- PMCID: PMC10722698
- DOI: 10.1186/s12987-023-00500-w
Structural characterization of SLYM-a 4th meningeal membrane
Abstract
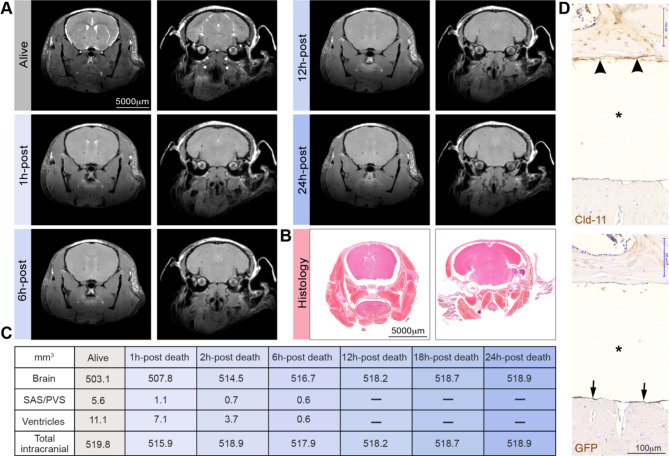
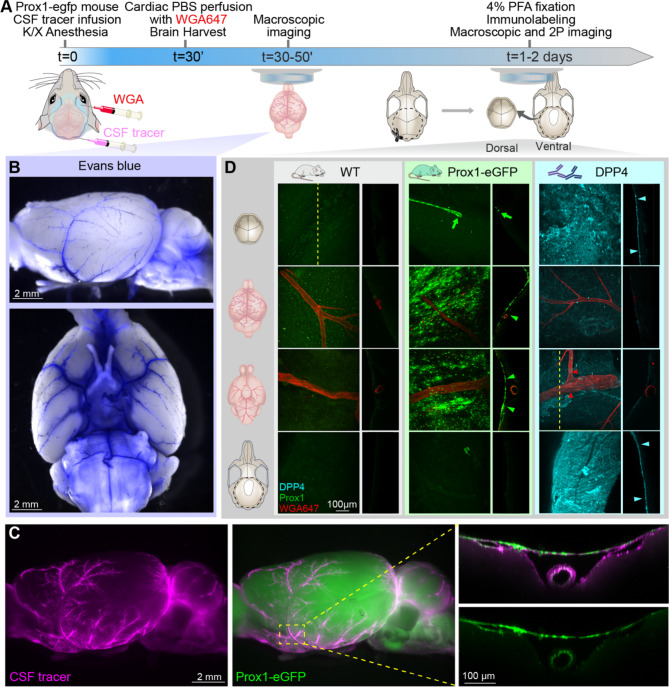
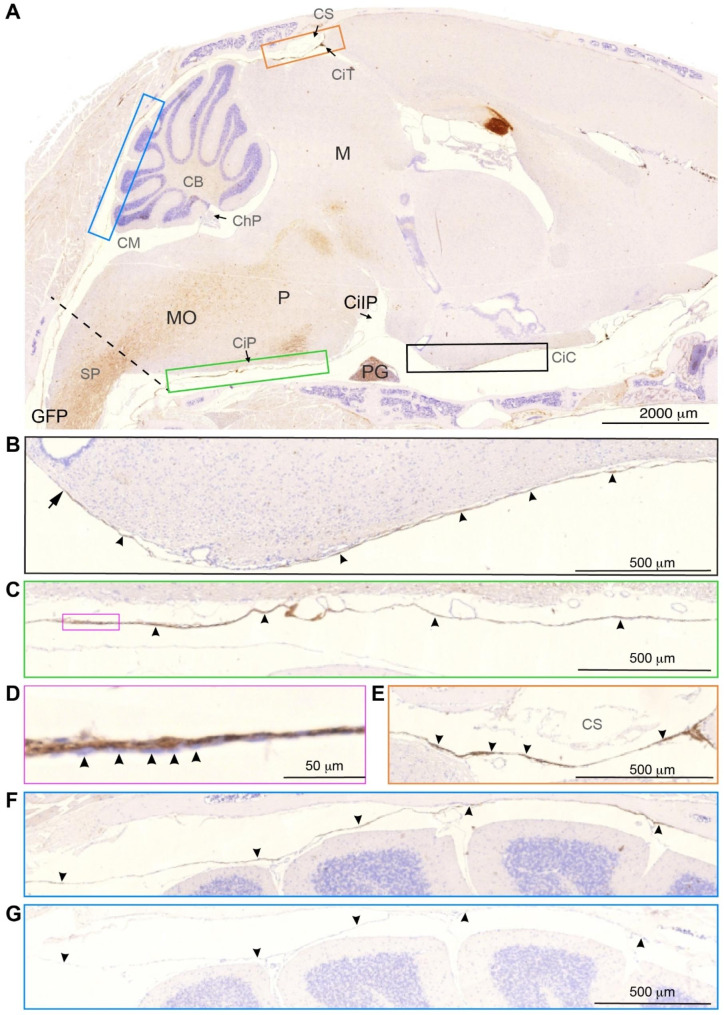
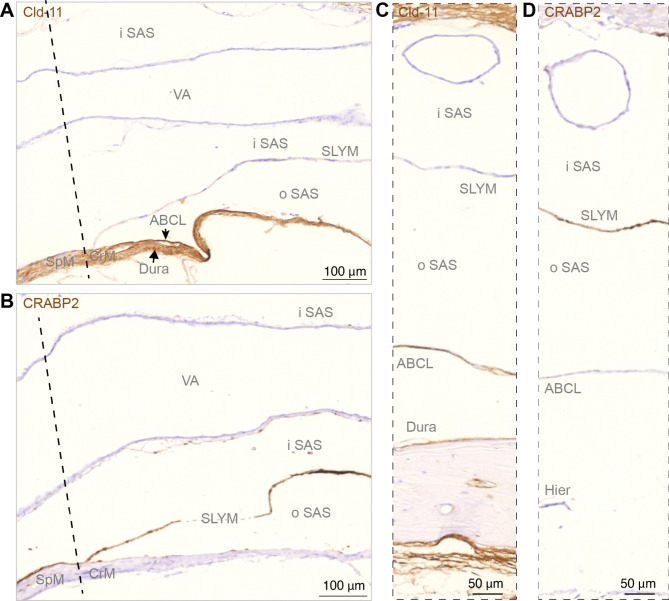
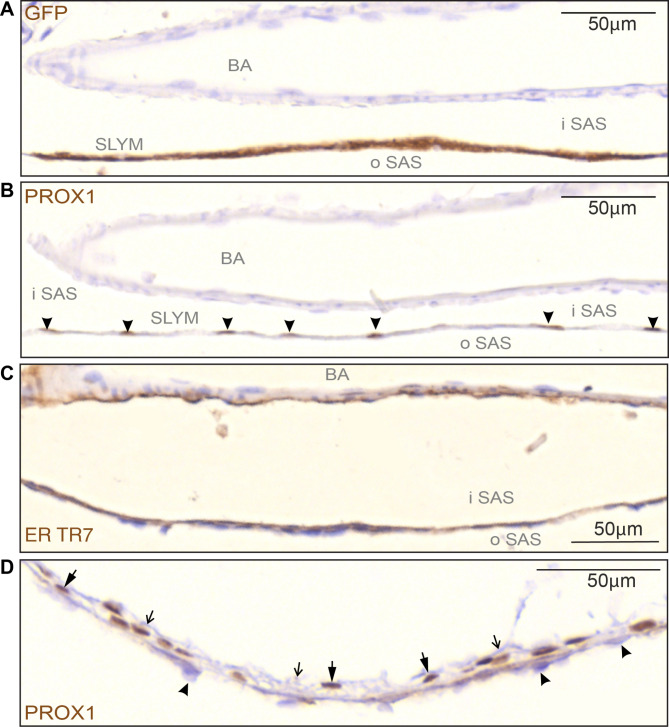
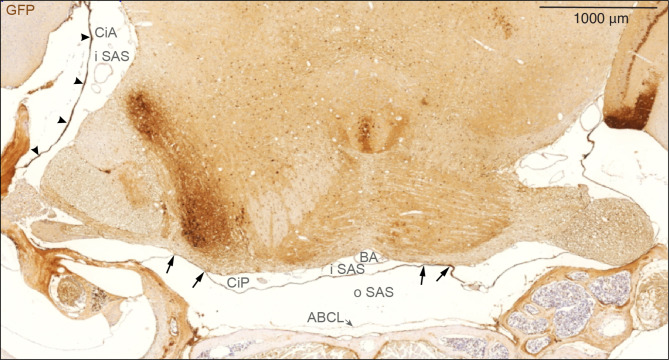
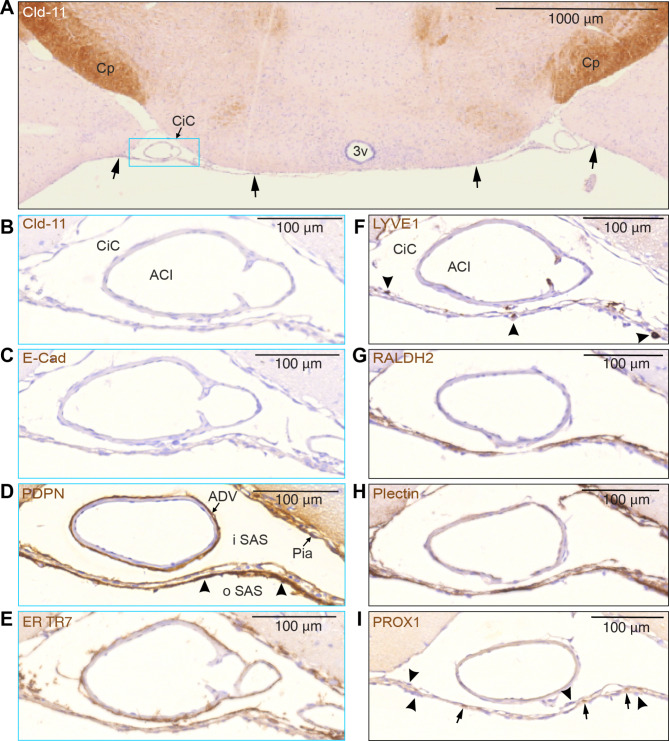
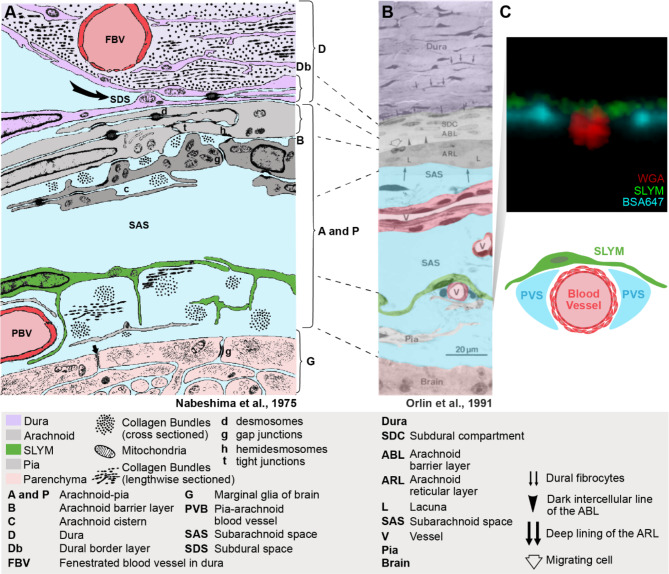
Traditionally, the meninges are described as 3 distinct layers, dura, arachnoid and pia. Yet, the classification of the connective meningeal membranes surrounding the brain is based on postmortem macroscopic examination. Ultrastructural and single cell transcriptome analyses have documented that the 3 meningeal layers can be subdivided into several distinct layers based on cellular characteristics. We here re-examined the existence of a 4th meningeal membrane, Subarachnoid Lymphatic-like Membrane or SLYM in Prox1-eGFP reporter mice. Imaging of freshly resected whole brains showed that SLYM covers the entire brain and brain stem and forms a roof shielding the subarachnoid cerebrospinal fluid (CSF)-filled cisterns and the pia-adjacent vasculature. Thus, SLYM is strategically positioned to facilitate periarterial influx of freshly produced CSF and thereby support unidirectional glymphatic CSF transport. Histological analysis showed that, in spinal cord and parts of dorsal cortex, SLYM fused with the arachnoid barrier layer, while in the basal brain stem typically formed a 1-3 cell layered membrane subdividing the subarachnoid space into two compartments. However, great care should be taken when interpreting the organization of the delicate leptomeningeal membranes in tissue sections. We show that hyperosmotic fixatives dehydrate the tissue with the risk of shrinkage and dislocation of these fragile membranes in postmortem preparations.
Keywords: Cerebrospinal fluid; Glymphatic system; Perivascular space; Prospero Homeobox 1; Subarachnoid space.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Structural characterization of SLYM - a 4th meningeal membrane.Res Sq [Preprint]. 2023 Oct 31:rs.3.rs-3500436. doi: 10.21203/rs.3.rs-3500436/v1. Res Sq. 2023. Update in: Fluids Barriers CNS. 2023 Dec 14;20(1):93. doi: 10.1186/s12987-023-00500-w. PMID: 37961391 Free PMC article. Updated. Preprint.
-
Structural characterization of SLYM - a 4 th meningeal membrane.bioRxiv [Preprint]. 2023 Oct 28:2023.10.20.563351. doi: 10.1101/2023.10.20.563351. bioRxiv. 2023. Update in: Fluids Barriers CNS. 2023 Dec 14;20(1):93. doi: 10.1186/s12987-023-00500-w. PMID: 37961461 Free PMC article. Updated. Preprint.
Comment in
-
An "outer subarachnoid space": fact or artifact? A commentary on "Structural characterization of SLYM- a 4th meningeal membrane" fluids and barriers of the CNS (2023) 20:93 by V. Plá et al.Fluids Barriers CNS. 2024 Jun 3;21(1):48. doi: 10.1186/s12987-024-00539-3. Fluids Barriers CNS. 2024. PMID: 38831302 Free PMC article. No abstract available.
-
Commentary on "Structural characterization of SLYM - a 4th meningeal membrane".Fluids Barriers CNS. 2024 Sep 9;21(1):69. doi: 10.1186/s12987-024-00568-y. Fluids Barriers CNS. 2024. PMID: 39252039 Free PMC article.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
