

AMPK-HIF-1α signaling enhances glucose-derived de novo serine biosynthesis to promote glioblastoma growth
- PMID: 38098117
- PMCID: PMC10722853
- DOI: 10.1186/s13046-023-02927-3
AMPK-HIF-1α signaling enhances glucose-derived de novo serine biosynthesis to promote glioblastoma growth
Abstract
Background: Cancer cells undergo cellular adaptation through metabolic reprogramming to sustain survival and rapid growth under various stress conditions. However, how brain tumors modulate their metabolic flexibility in the naturally serine/glycine (S/G)-deficient brain microenvironment remain unknown.
Methods: We used a range of primary/stem-like and established glioblastoma (GBM) cell models in vitro and in vivo. To identify the regulatory mechanisms of S/G deprivation-induced metabolic flexibility, we employed high-throughput RNA-sequencing, transcriptomic analysis, metabolic flux analysis, metabolites analysis, chromatin immunoprecipitation (ChIP), luciferase reporter, nuclear fractionation, cycloheximide-chase, and glucose consumption. The clinical significances were analyzed in the genomic database (GSE4290) and in human GBM specimens.
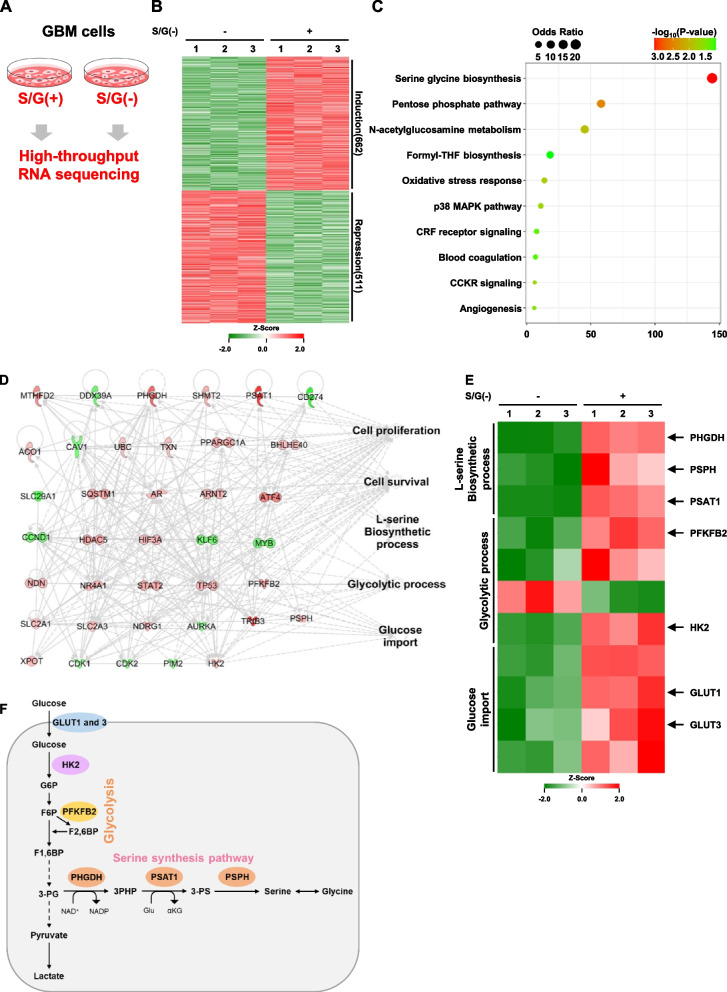
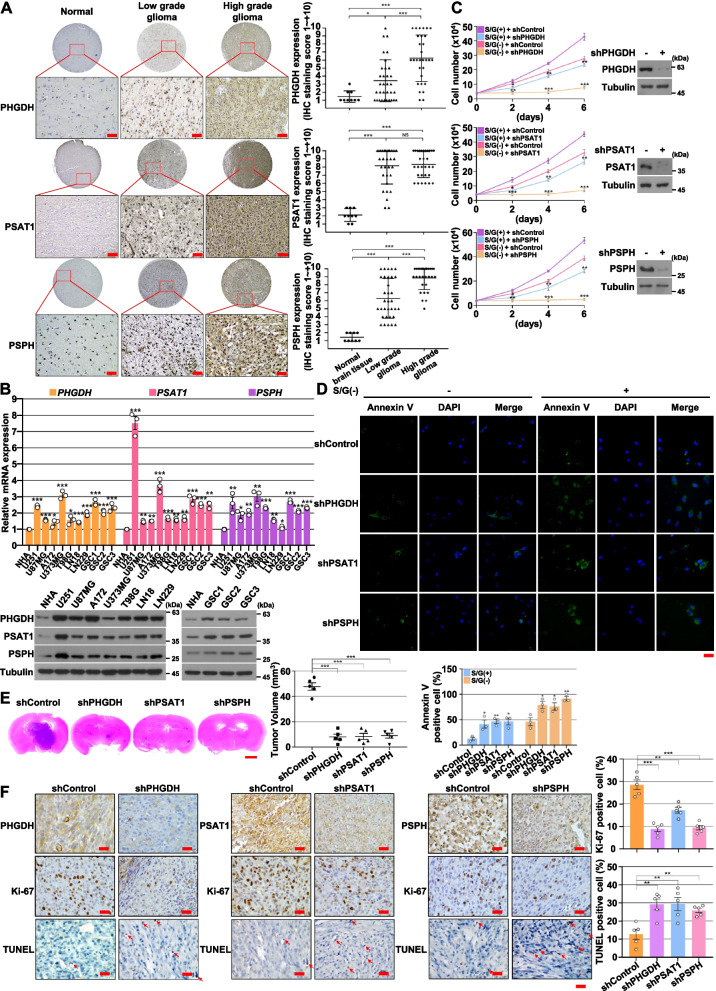
Results: The high-throughput RNA-sequencing and transcriptomic analysis demonstrate that the de novo serine synthesis pathway (SSP) and glycolysis are highly activated in GBM cells under S/G deprivation conditions. Mechanistically, S/G deprivation rapidly induces reactive oxygen species (ROS)-mediated AMP-activated protein kinase (AMPK) activation and AMPK-dependent hypoxia-inducible factor (HIF)-1α stabilization and transactivation. Activated HIF-1α in turn promotes the expression of SSP enzymes phosphoglycerate dehydrogenase (PHGDH), phosphoserine aminotransferase 1 (PSAT1), and phosphoserine phosphatase (PSPH). In addition, the HIF-1α-induced expression of glycolytic genes (GLUT1, GLUT3, HK2, and PFKFB2) promotes glucose uptake, glycolysis, and glycolytic flux to fuel SSP, leading to elevated de novo serine and glycine biosynthesis, NADPH/NADP+ ratio, and the proliferation and survival of GBM cells. Analyses of human GBM specimens reveal that the levels of overexpressed PHGDH, PSAT1, and PSPH are positively correlated with levels of AMPK T172 phosphorylation and HIF-1α expression and the poor prognosis of GBM patients.
Conclusion: Our findings reveal that metabolic stress-enhanced glucose-derived de novo serine biosynthesis is a critical metabolic feature of GBM cells, and highlight the potential to target SSP for treating human GBM.
Keywords: AMPK; De novo serine synthesis; Glycine; HIF-1α; Serine.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
MeSH terms
Substances
Grants and funding
- RS-2023-00208431/National research foundation of korea
- 82188102/National natural science foundation of china
- 82030074/National natural science foundation of china
- 2020YFA0803300/Ministry of science and technology of the people's republic of china
- NCTIB2022HS02006/National center of technology innovation for biopharmaceuticals
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
