

Mechanisms of aquaporin-4 vesicular trafficking in mammalian cells
- PMID: 38102893
- PMCID: PMC10953025
- DOI: 10.1111/jnc.16029
Mechanisms of aquaporin-4 vesicular trafficking in mammalian cells
Abstract
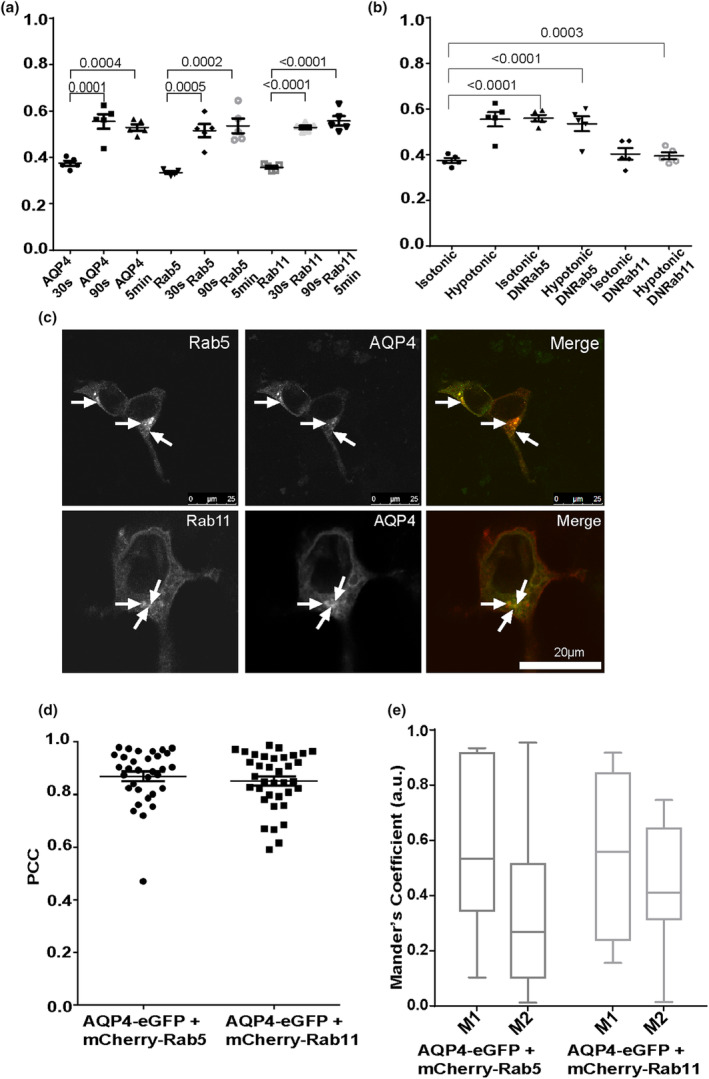
The aquaporin-4 (AQP4) water channel is abundantly expressed in the glial cells of the central nervous system and facilitates brain swelling following diverse insults, such as traumatic injury or stroke. Lack of specific and therapeutic AQP4 inhibitors highlights the need to explore alternative routes to control the water permeability of glial cell membranes. The cell surface abundance of AQP4 in mammalian cells fluctuates rapidly in response to changes in oxygen levels and tonicity, suggesting a role for vesicular trafficking in its translocation to and from the cell surface. However, the molecular mechanisms of AQP4 trafficking are not fully elucidated. In this work, early and recycling endosomes were investigated as likely candidates of rapid AQP4 translocation together with changes in cytoskeletal dynamics. In transiently transfected HEK293 cells a significant amount of AQP-eGFP colocalised with mCherry-Rab5-positive early endosomes and mCherry-Rab11-positive recycling endosomes. When exposed to hypotonic conditions, AQP4-eGFP rapidly translocated from intracellular vesicles to the cell surface. Co-expression of dominant negative forms of the mCherry-Rab5 and -Rab11 with AQP4-eGFP prevented hypotonicity-induced AQP4-eGFP trafficking and led to concentration at the cell surface or intracellular vesicles respectively. Use of endocytosis inhibiting drugs indicated that AQP4 internalisation was dynamin-dependent. Cytoskeleton dynamics-modifying drugs also affected AQP4 translocation to and from the cell surface. AQP4 trafficking mechanisms were validated in primary human astrocytes, which express high levels of endogenous AQP4. The results highlight the role of early and recycling endosomes and cytoskeletal dynamics in AQP4 translocation in response to hypotonic and hypoxic stress and suggest continuous cycling of AQP4 between intracellular vesicles and the cell surface under physiological conditions.
Keywords: Rab GTPase; aquaporin-4; astrocyte; cytoskeleton; oedema; vesicular trafficking.
© 2023 The Authors. Journal of Neurochemistry published by John Wiley & Sons Ltd on behalf of International Society for Neurochemistry.
Conflict of interest statement
R.M.B., P.K. and M.M.S. are shareholders in Estuar Pharmaceuticals.
Figures





References
-
- Badaut, J. , Lasbennes, F. , Magistretti, P. J. , & Regli, L. (2002). Aquaporins in brain: Distribution, physiology, and pathophysiology. Journal of Cerebral Blood Flow and Metabolism, 22(4), 367–378. - PubMed
-
- Carmosino, M. , Procino, G. , Tamma, G. , Mannucci, R. , Svelto, M. , & Valenti, G. (2007). Trafficking and phosphorylation dynamics of AQP4 in histamine‐treated human gastric cells. Biology of the Cell, 99(1), 25–36. - PubMed
-
- Casella, J. F. , Flanagan, M. D. , & Lin, S. (1981). Cytochalasin D inhibits Actin polymerization and induces depolymerization of Actin filaments formed during platelet shape change. Nature, 293(5830), 302–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
