

Metabolic reprogramming by Syntenin-1 directs RA FLS and endothelial cell-mediated inflammation and angiogenesis
- PMID: 38105293
- PMCID: PMC10757714
- DOI: 10.1038/s41423-023-01108-8
Metabolic reprogramming by Syntenin-1 directs RA FLS and endothelial cell-mediated inflammation and angiogenesis
Abstract
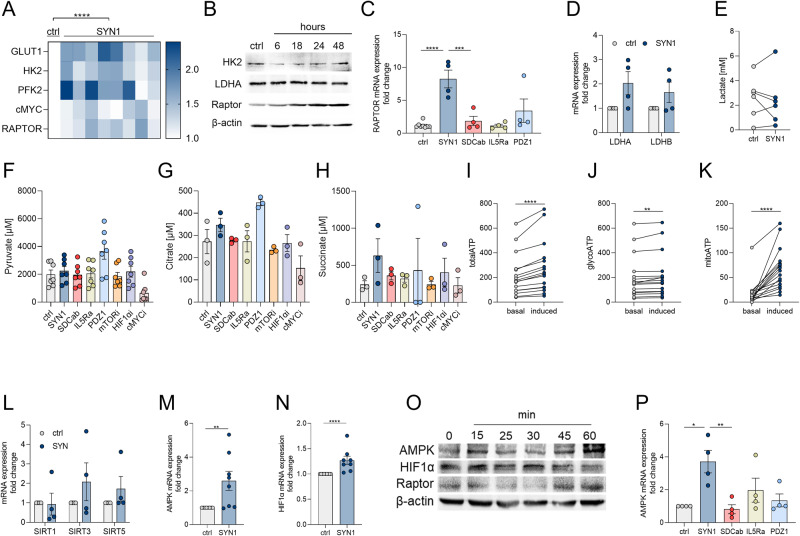
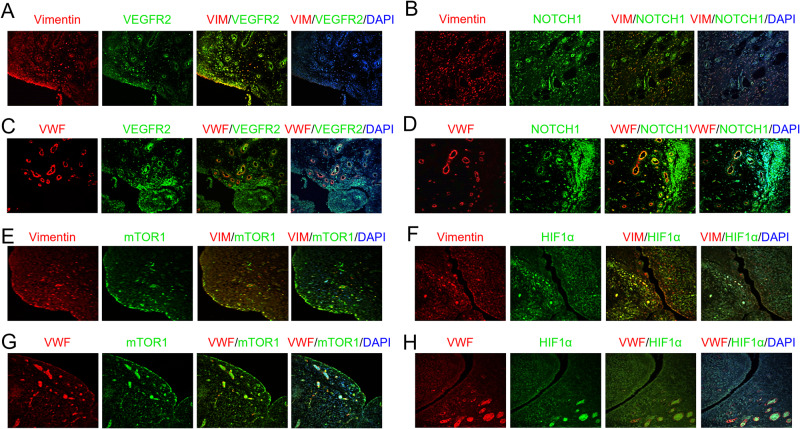
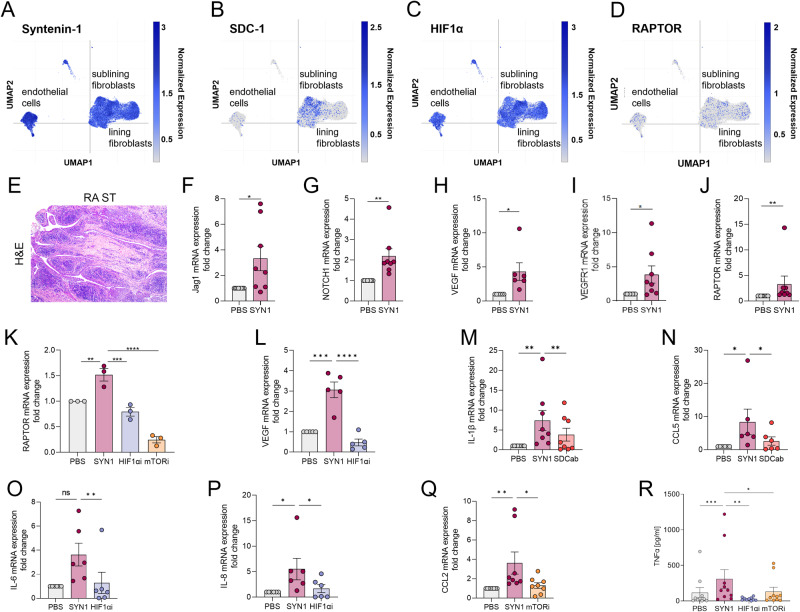
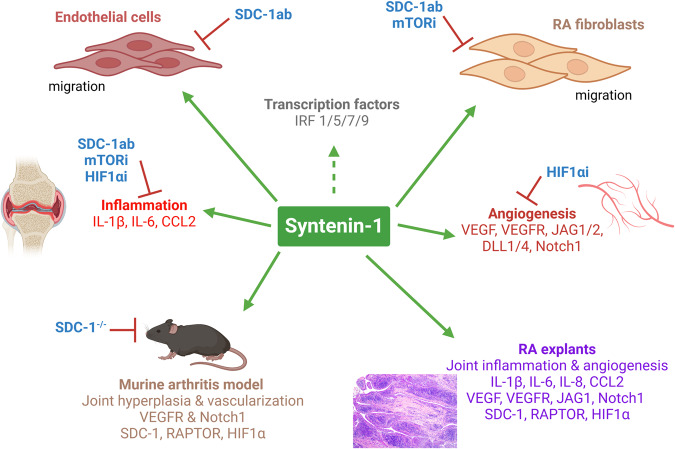
A novel rheumatoid arthritis (RA) synovial fluid protein, Syntenin-1, and its receptor, Syndecan-1 (SDC-1), are colocalized on RA synovial tissue endothelial cells and fibroblast-like synoviocytes (FLS). Syntenin-1 exacerbates the inflammatory landscape of endothelial cells and RA FLS by upregulating transcription of IRF1/5/7/9, IL-1β, IL-6, and CCL2 through SDC-1 ligation and HIF1α, or mTOR activation. Mechanistically, Syntenin-1 orchestrates RA FLS and endothelial cell invasion via SDC-1 and/or mTOR signaling. In Syntenin-1 reprogrammed endothelial cells, the dynamic expression of metabolic intermediates coincides with escalated glycolysis along with unchanged oxidative factors, AMPK, PGC-1α, citrate, and inactive oxidative phosphorylation. Conversely, RA FLS rewired by Syntenin-1 displayed a modest glycolytic-ATP accompanied by a robust mitochondrial-ATP capacity. The enriched mitochondrial-ATP detected in Syntenin-1 reprogrammed RA FLS was coupled with mitochondrial fusion and fission recapitulated by escalated Mitofusin-2 and DRP1 expression. We found that VEGFR1/2 and Notch1 networks are responsible for the crosstalk between Syntenin-1 rewired endothelial cells and RA FLS, which are also represented in RA explants. Similar to RA explants, morphological and transcriptome studies authenticated the importance of VEGFR1/2, Notch1, RAPTOR, and HIF1α pathways in Syntenin-1 arthritic mice and their obstruction in SDC-1 deficient animals. Consistently, dysregulation of SDC-1, mTOR, and HIF1α negated Syntenin-1 inflammatory phenotype in RA explants, while inhibition of HIF1α impaired synovial angiogenic imprint amplified by Syntenin-1. In conclusion, since the current therapies are ineffective on Syntenin-1 and SDC-1 expression in RA synovial tissue and blood, targeting this pathway and its interconnected metabolic intermediates may provide a novel therapeutic strategy.
Keywords: RA FLS; RA explants; Syndecan-1; Syntenin-1; immunometabolism.
© 2023. The Author(s), under exclusive licence to CSI and USTC.
Conflict of interest statement
The authors declare no competing interests.
Figures
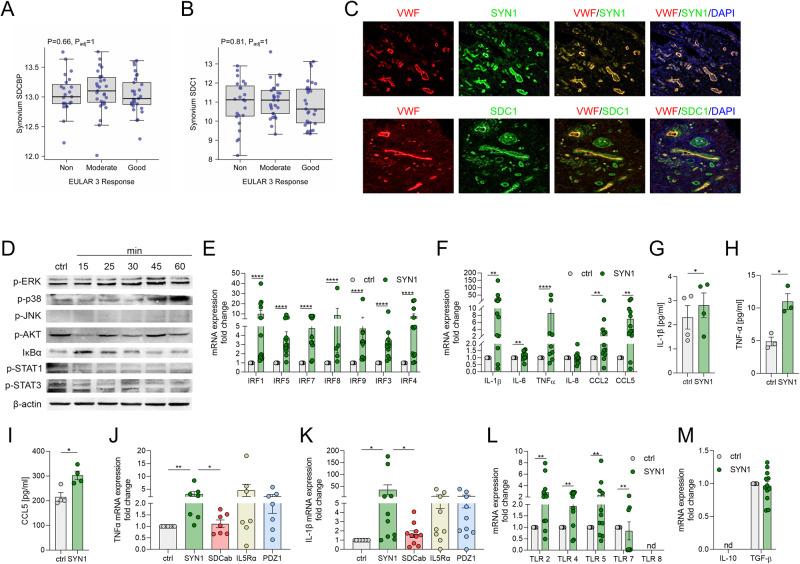
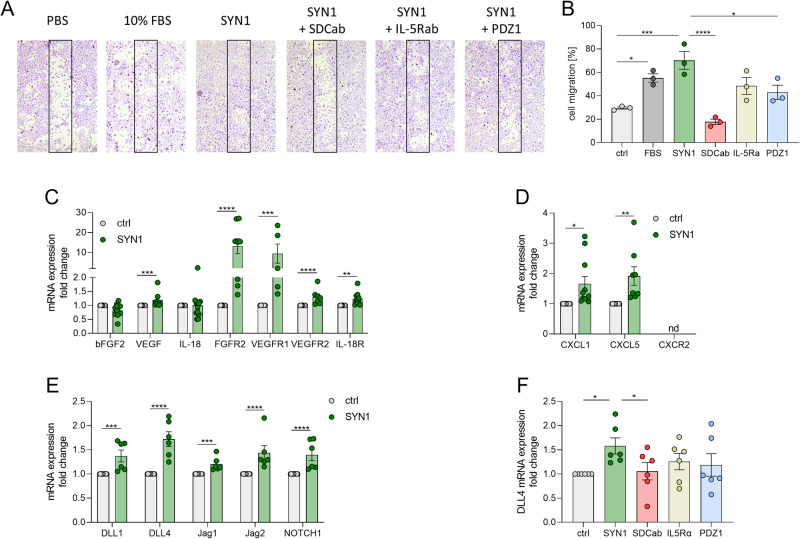
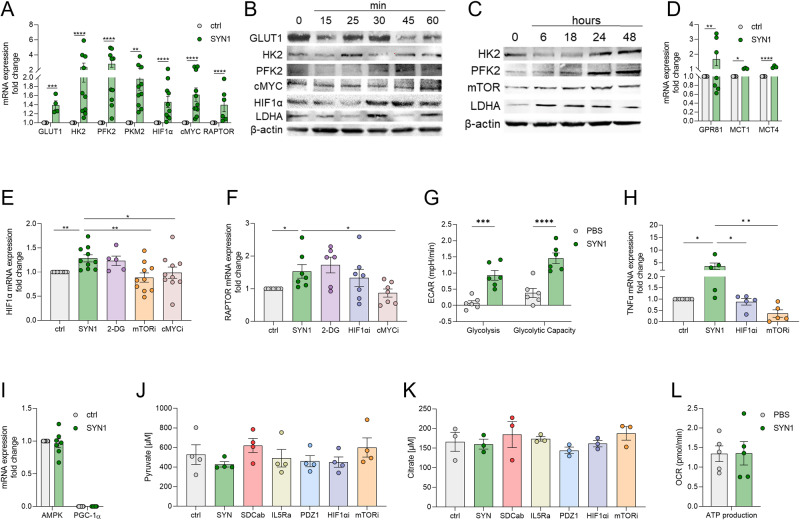
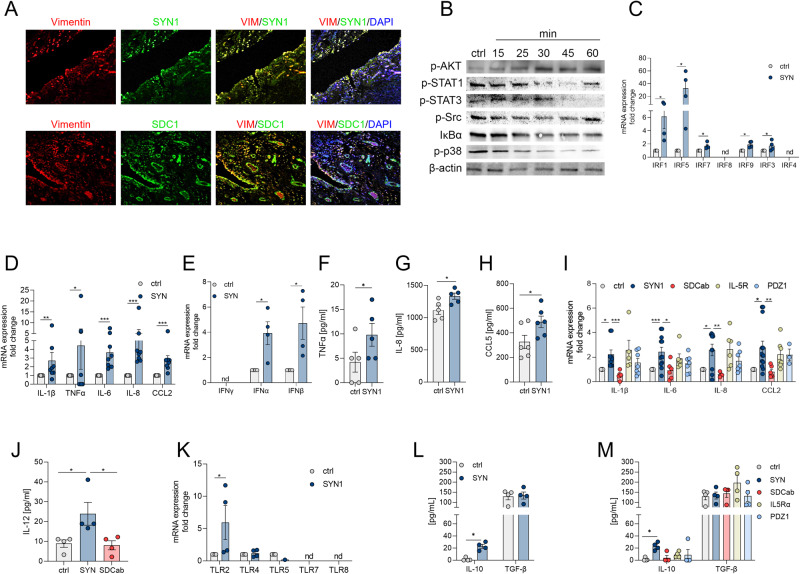


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
