

This is a preprint.
Electrostatic encoding of genome organization principles within single native nucleosomes
- PMID: 38106048
- PMCID: PMC10723453
- DOI: 10.1101/2023.12.08.570828
Electrostatic encoding of genome organization principles within single native nucleosomes
Update in
-
Native nucleosomes intrinsically encode genome organization principles.Nature. 2025 Jul;643(8071):572-581. doi: 10.1038/s41586-025-08971-7. Epub 2025 May 7. Nature. 2025. PMID: 40335690 Free PMC article.
Abstract
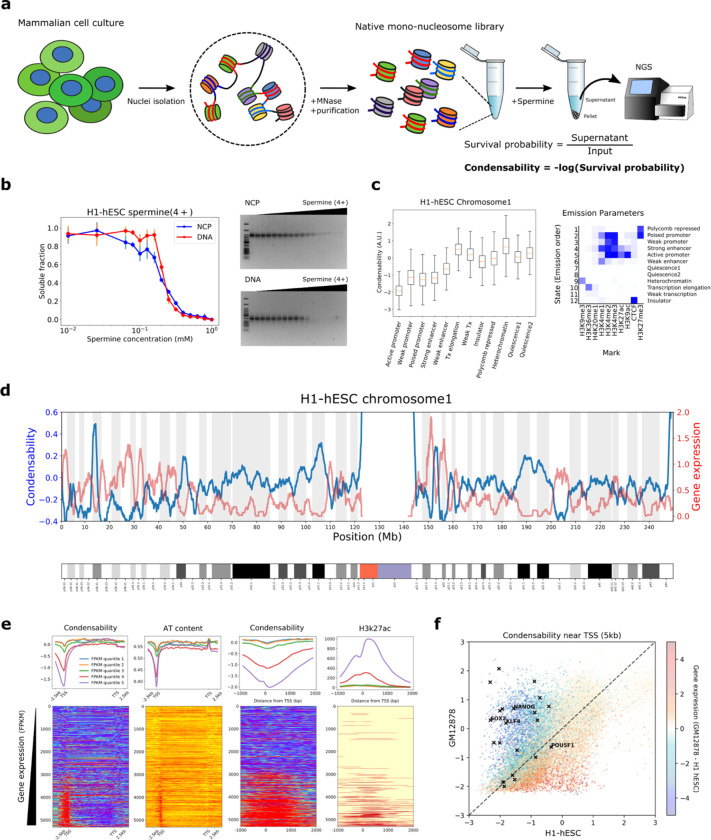
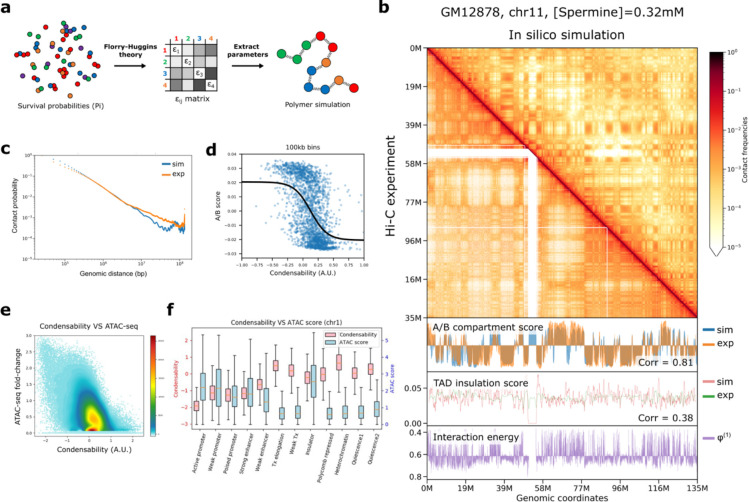
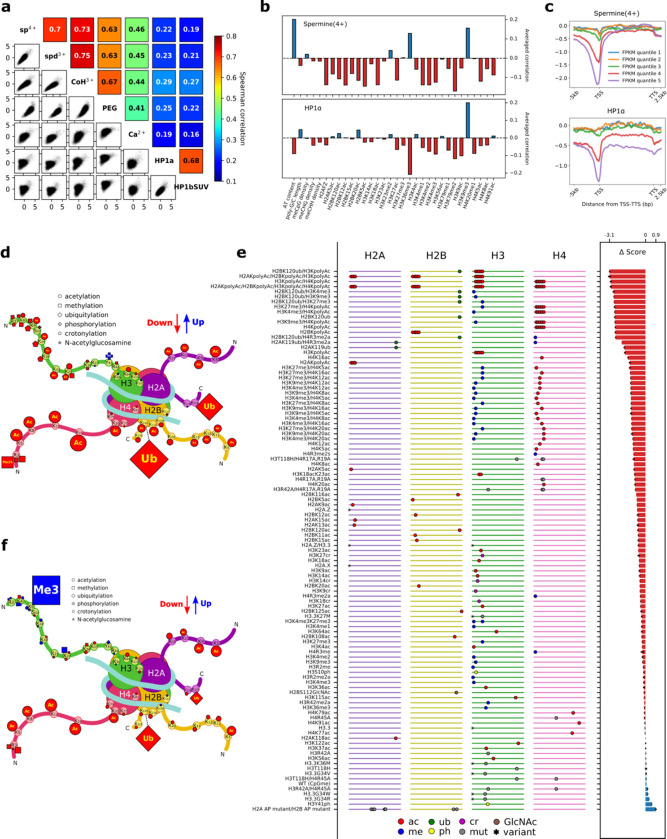
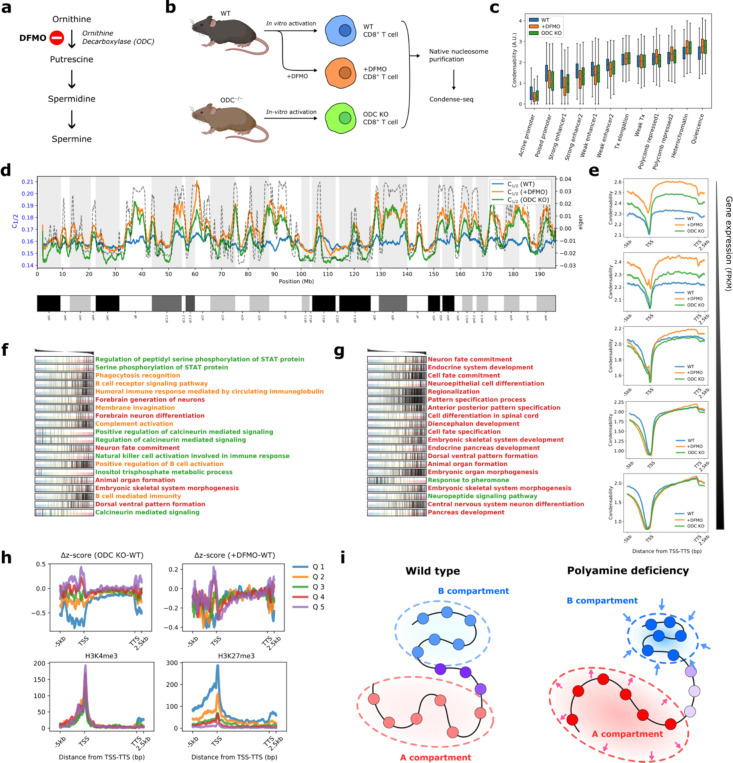
The eukaryotic genome, first packed into nucleosomes of about 150 bp around the histone core, is organized into euchromatin and heterochromatin, corresponding to the A and B compartments, respectively. Here, we asked if individual nucleosomes in vivo know where to go. That is, do mono-nucleosomes by themselves contain A/B compartment information, associated with transcription activity, in their biophysical properties? We purified native mono-nucleosomes to high monodispersity and used physiological concentrations of biological polyamines to determine their condensability. The chromosomal regions known to partition into A compartments have low condensability and vice versa. In silico chromatin polymer simulations using condensability as the only input showed that biophysical information needed to form compartments is all contained in single native nucleosomes and no other factors are needed. Condensability is also strongly anticorrelated with gene expression, and especially so near the promoter region and in a cell type dependent manner. Therefore, individual nucleosomes in the promoter know whether the gene is on or off, and that information is contained in their biophysical properties. Comparison with genetic and epigenetic features suggest that nucleosome condensability is a very meaningful axis onto which to project the high dimensional cellular chromatin state. Analysis of condensability using various condensing agents including those that are protein-based suggests that genome organization principle encoded into individual nucleosomes is electrostatic in nature. Polyamine depletion in mouse T cells, by either knocking out ornithine decarboxylase (ODC) or inhibiting ODC, results in hyperpolarized condensability, suggesting that when cells cannot rely on polyamines to translate biophysical properties of nucleosomes to control gene expression and 3D genome organization, they accentuate condensability contrast, which may explain dysfunction known to occur with polyamine deficiency.
Conflict of interest statement
CONFLICT OF INTEREST The authors declare no conflicts of interest.
Figures
References
-
- Kempfer R. & Pombo A. Methods for mapping 3D chromosome architecture. Nature Reviews Genetics 2019 21:4 21, 207–226 (2019). - PubMed
-
- Zheng H. & Xie W. The role of 3D genome organization in development and cell differentiation. Nature Reviews Molecular Cell Biology 2019 20:9 20, 535–550 (2019). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials