

This is a preprint.
Conserved autism-associated genes tune social feeding behavior in C. elegans
- PMID: 38106124
- PMCID: PMC10723370
- DOI: 10.1101/2023.12.05.570116
Conserved autism-associated genes tune social feeding behavior in C. elegans
Update in
-
Conserved autism-associated genes tune social feeding behavior in C. elegans.Nat Commun. 2024 Oct 28;15(1):9301. doi: 10.1038/s41467-024-53590-x. Nat Commun. 2024. PMID: 39468047 Free PMC article.
Abstract
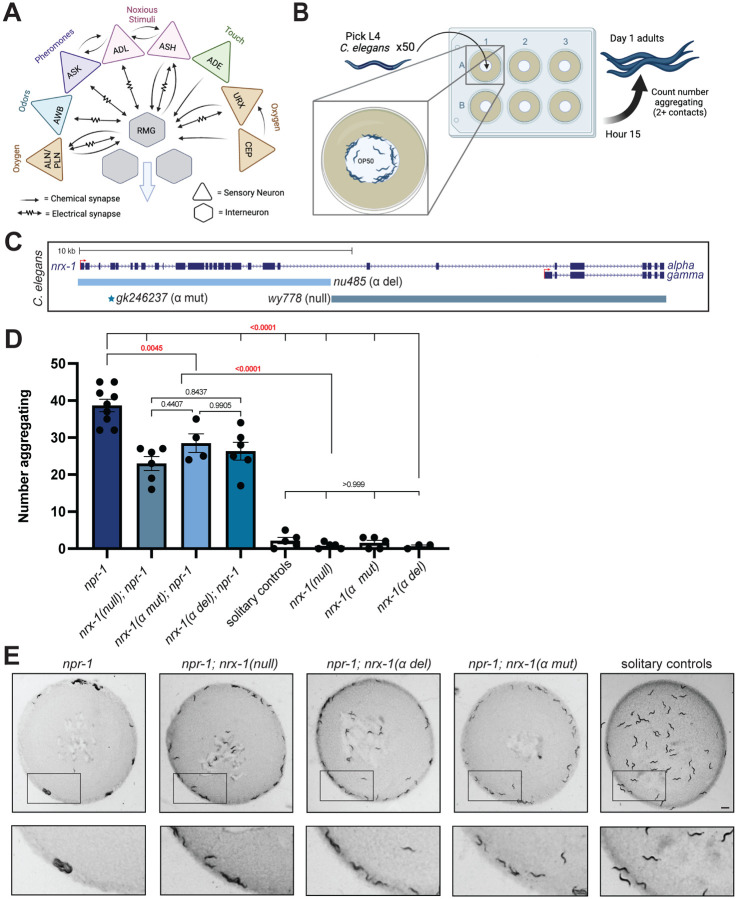
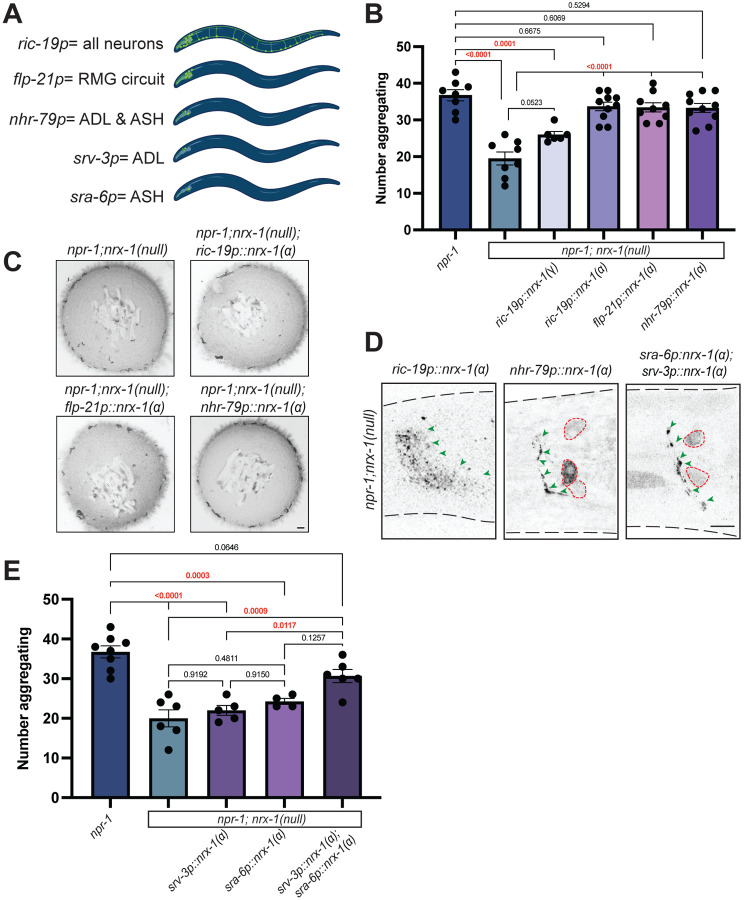
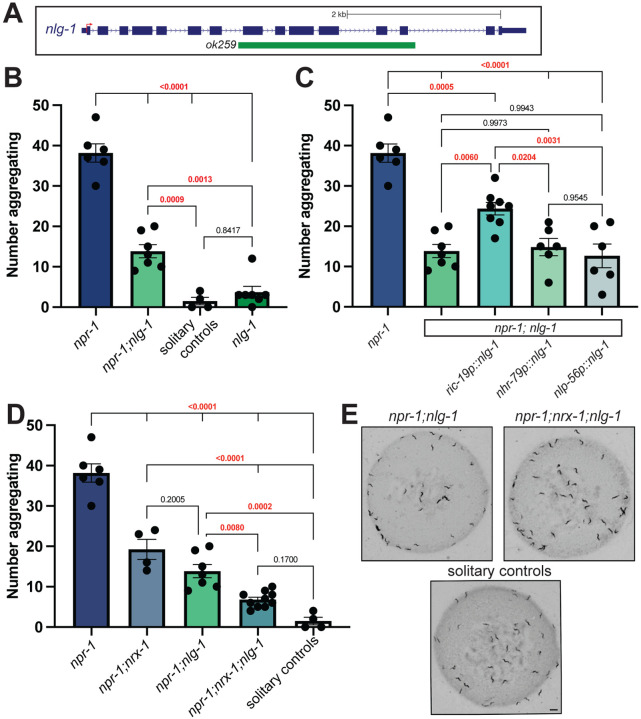
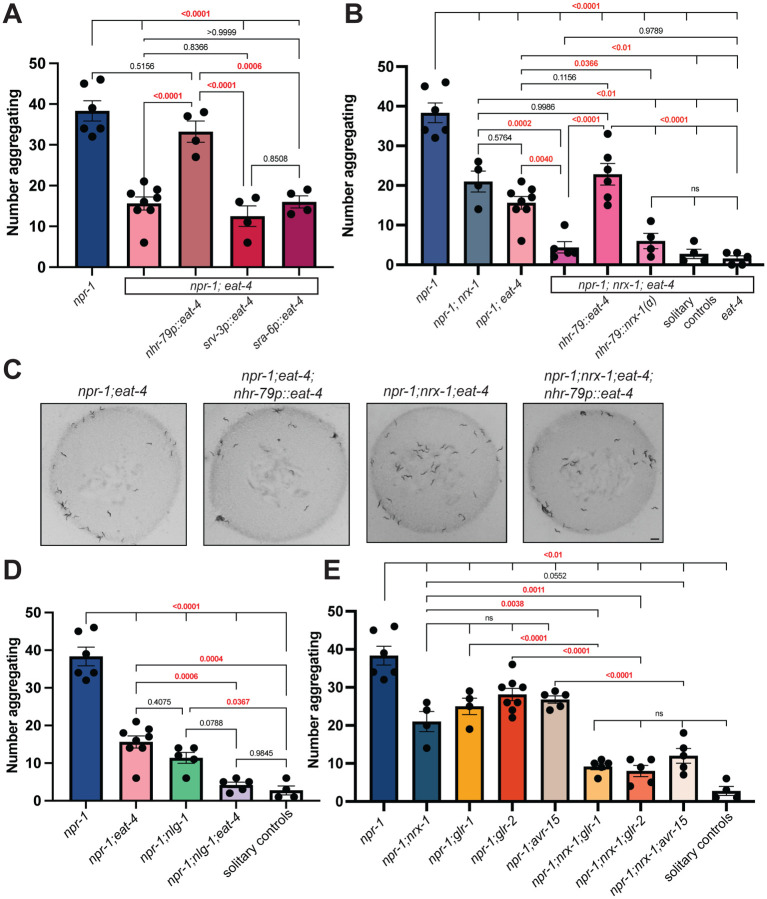
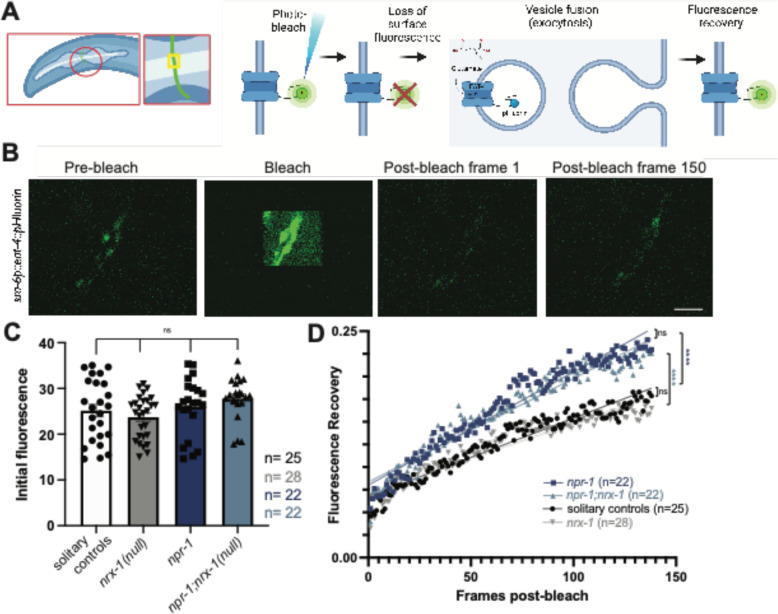
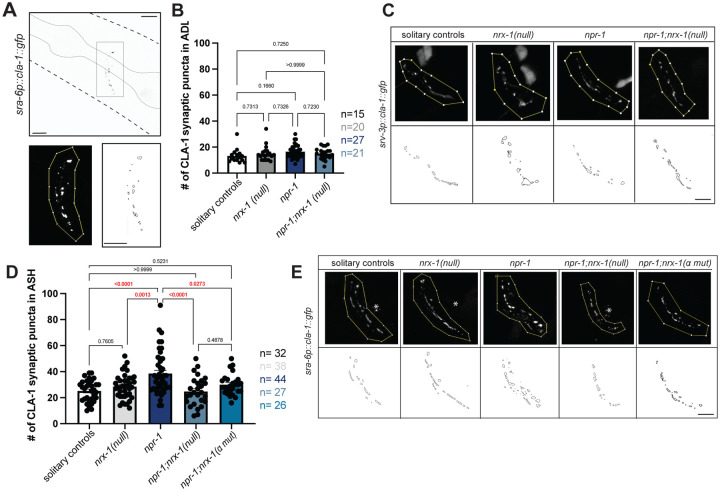
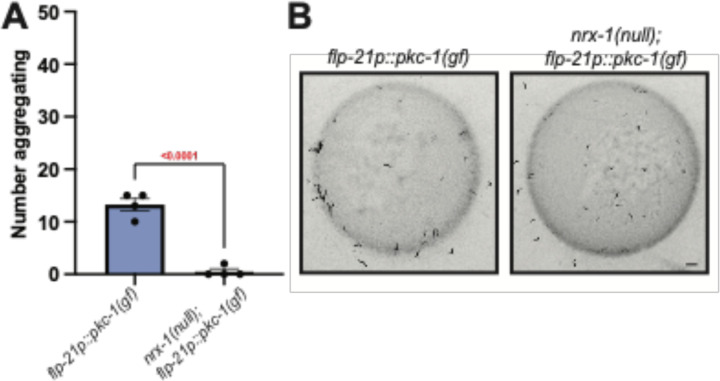
Animal foraging is an essential and evolutionarily conserved behavior that occurs in social and solitary contexts, but the underlying molecular pathways are not well defined. We discover that conserved autism-associated genes (NRXN1(nrx-1), NLGN3(nlg-1), GRIA1,2,3(glr-1), GRIA2(glr-2), and GLRA2,GABRA3(avr-15)) regulate aggregate feeding in C. elegans, a simple social behavior. NRX-1 functions in chemosensory neurons (ADL and ASH) independently of its postsynaptic partner NLG-1 to regulate social feeding. Glutamate from these neurons is also crucial for aggregate feeding, acting independently of NRX-1 and NLG-1. Compared to solitary counterparts, social animals show faster presynaptic release and more presynaptic release sites in ASH neurons, with only the latter requiring nrx-1. Disruption of these distinct signaling components additively converts behavior from social to solitary. Aggregation induced by circuit activation is also dependent on nrx-1. Collectively, we find that aggregate feeding is tuned by conserved autism-associated genes through complementary synaptic mechanisms, revealing molecular principles driving social feeding.
Conflict of interest statement
Competing Interests: The authors declare no conflicts of interest.
Figures
References
-
- Davidson JD, Arauco-Aliaga RP, Crow S, Gordon DM, Goldman MS. Effect of Interactions between Harvester Ants on Forager Decisions. Frontiers in Ecology and Evolution. 2016;4. Accessed October 9, 2023. https://www.frontiersin.org/articles/10.3389/fevo.2016.00115 - DOI - PMC - PubMed
-
- Gordon DM. Ant Encounters: Interaction Networks and Colony Behavior. In: Ant Encounters. Princeton University Press; 2010. doi:10.1515/9781400835447 - DOI
-
- Azorsa F, Muscedere ML, Traniello JFA. Socioecology and Evolutionary Neurobiology of Predatory Ants. Frontiers in Ecology and Evolution. 2022;9. Accessed October 9, 2023. https://www.frontiersin.org/articles/10.3389/fevo.2021.804200 - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous