

Ca2+-regulated expression of high affinity methylaminoisobutryic acid transport in hippocampal neurons inhibited by riluzole and novel neuroprotective aminothiazoles
- PMID: 38107787
- PMCID: PMC10724208
- DOI: 10.1016/j.crphys.2023.100109
Ca2+-regulated expression of high affinity methylaminoisobutryic acid transport in hippocampal neurons inhibited by riluzole and novel neuroprotective aminothiazoles
Abstract
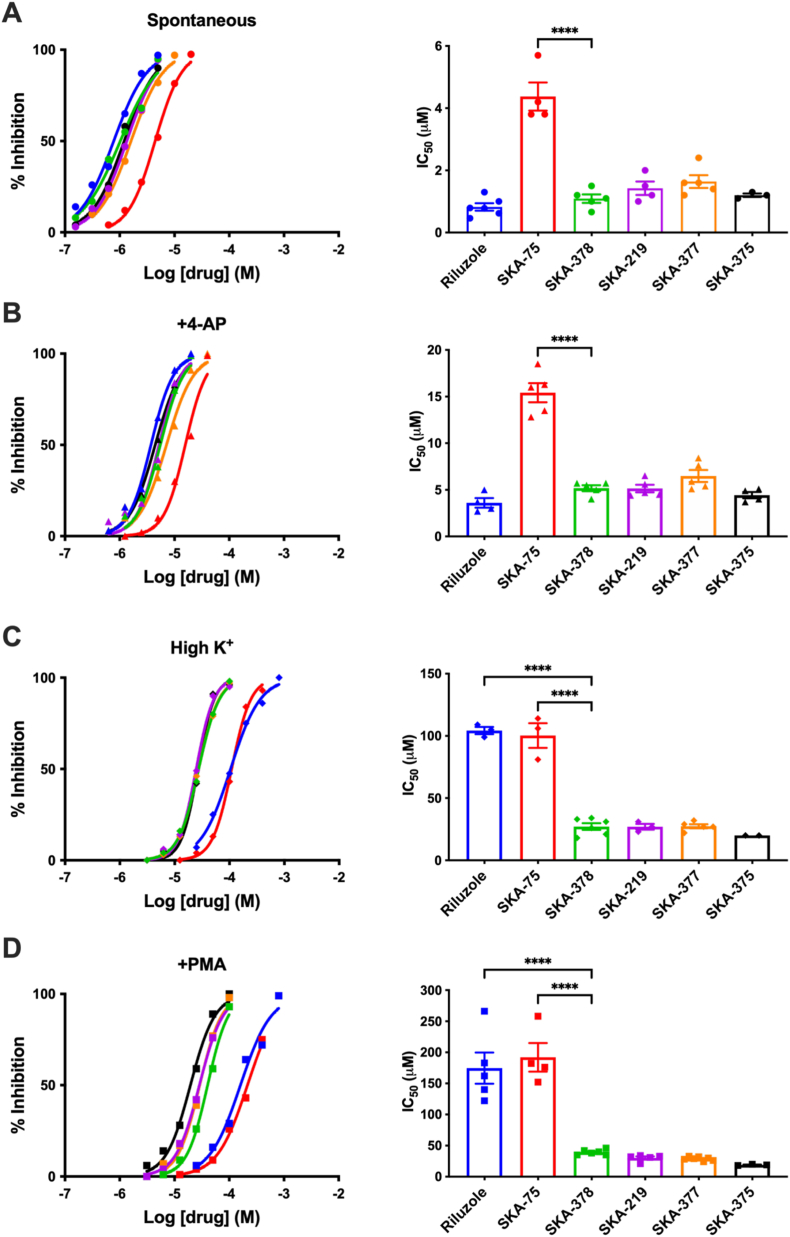
High affinity methylaminoisobutyric acid(MeAIB)/glutamine(Gln) transport activity regulated by neuronal firing occurs at the plasma membrane in mature rat hippocampal neuron-enriched cultures. Spontaneous Ca2+-regulated transport activity was similarly inhibited by riluzole, a benzothiazole anticonvulsant agent, and by novel naphthalenyl substituted aminothiazole derivatives such as SKA-378. Here, we report that spontaneous transport activity is stimulated by 4-aminopyridine (4-AP) and that phorbol-myristate acetate (PMA) increases high K+ stimulated transport activity that is inhibited by staurosporine. 4-AP-stimulated spontaneous and PMA-stimulated high K+-induced transport is not present at 7 days in vitro (DIV) and is maximal by DIV∼21. The relative affinity for MeAIB is similar for spontaneous and high K+-stimulated transport (Km ∼ 50 μM) suggesting that a single transporter is involved. While riluzole and SKA-378 inhibit spontaneous transport with equal potency (IC50 ∼ 1 μM), they exhibit decreased (∼3-5 X) potency for 4-AP-stimulated spontaneous transport. Interestingly, high K+-stimulated MeAIB transport displays lower and differential sensitivity to the two compounds. SKA-378-related halogenated derivatives of SKA-75 (SKA-219, SKA-377 and SKA-375) preferentially inhibit high K+-induced expression of MeAIB transport activity at the plasma membrane (IC50 < 25 μM), compared to SKA-75 and riluzole (IC50 > 100 μM). Ca2+-dependent spontaneous and high K+-stimulated MeAIB transport activity is blocked by ω-conotoxin MVIIC, ω-agatoxin IVA, ω-agatoxin TK (IC50 ∼ 500 nM) or cadmium ion (IC50 ∼ 20 μM) demonstrating that P/Q-type CaV channels that are required for activity-regulated presynaptic vesicular glutamate (Glu) release are also required for high-affinity MeAIB transport expression at the plasma membrane. We suggest that neural activity driven and Ca2+ dependent trafficking of the high affinity MeAIB transporter to the plasma membrane is a unique target to understand mechanisms of Glu/Gln recycling in synapses and acute neuroprotection against excitotoxic presynaptic Glu induced neural injury.
Keywords: Glutamate/glutamine cycle; Hippocampal neurons; Methylaminoisobutyric acid/glutamine transporter; Riluzole; SKA-378; Vesicle recycling.
© 2023 The Authors.
Conflict of interest statement
The authors have no conflict of interests.
Figures




References
-
- Albrecht J., Sonnewald U., Waagepetersen H.S., Schousboe A. Glutamine in the central nervous system: function and dysfunction. Front. Biosci. 2007;12:332–343. - PubMed
-
- Amara S.G., Fontana A.C. Excitatory amino acid transporters: keeping up with glutamate. Neurochem. Int. 2002;41:313–318. - PubMed
-
- Bae H.-J., Lee Y.-S., Kang D.-W., Gu J.-S., Yoon B.-W., Roh J.-K. Neuroprotective effect of low dose riluzole in gerbil model of transient global ischemia. Neurosci. Lett. 2000;294:29–32. - PubMed
-
- Boulland J.L., Qureshi T., Seal R.P., et al. Expression of the vesicular glutamate transporters during development indicates the widespread corelease of multiple neurotransmitters. J. Comp. Neurol. 2004;480:264–280. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
