

Exploring the conformational changes of the Munc18-1/syntaxin 1a complex
- PMID: 38109275
- PMCID: PMC10895456
- DOI: 10.1002/pro.4870
Exploring the conformational changes of the Munc18-1/syntaxin 1a complex
Abstract
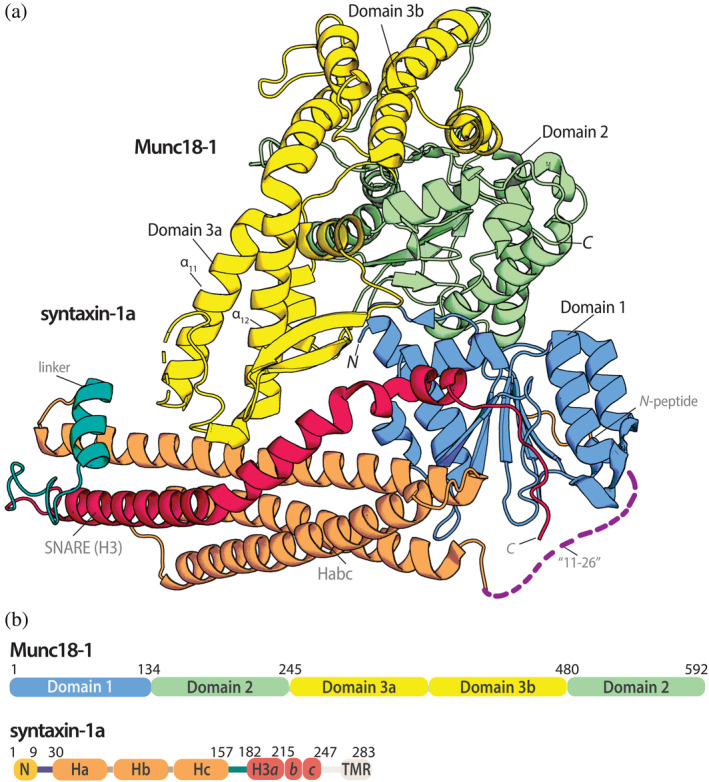
Neurotransmitters are released from synaptic vesicles, the membrane of which fuses with the plasma membrane upon calcium influx. This membrane fusion reaction is driven by the formation of a tight complex comprising the plasma membrane N-ethylmaleimide-sensitive factor attachment receptor (SNARE) proteins syntaxin-1a and SNAP-25 with the vesicle SNARE protein synaptobrevin. The neuronal protein Munc18-1 forms a stable complex with syntaxin-1a. Biochemically, syntaxin-1a cannot escape the tight grip of Munc18-1, so formation of the SNARE complex is inhibited. However, Munc18-1 is essential for the release of neurotransmitters in vivo. It has therefore been assumed that Munc18-1 makes the bound syntaxin-1a available for SNARE complex formation. Exactly how this occurs is still unclear, but it is assumed that structural rearrangements occur. Here, we used a series of mutations to specifically weaken the complex at different positions in order to induce these rearrangements biochemically. Our approach was guided through sequence and structural analysis and supported by molecular dynamics simulations. Subsequently, we created a homology model showing the complex in an altered conformation. This conformation presumably represents a more open arrangement of syntaxin-1a that permits the formation of a SNARE complex to be initiated while still bound to Munc18-1. In the future, research should investigate how this central reaction for neuronal communication is controlled by other proteins.
Keywords: Munc13; Munc18-1; SM protein; SNARE protein; neuronal secretion; synaptobrevin; syntaxin-1a.
© 2023 The Authors. Protein Science published by Wiley Periodicals LLC on behalf of The Protein Society.
Conflict of interest statement
The authors declare no competing interests.
Figures





Similar articles
-
Conformational change of syntaxin linker region induced by Munc13s initiates SNARE complex formation in synaptic exocytosis.EMBO J. 2017 Mar 15;36(6):816-829. doi: 10.15252/embj.201695775. Epub 2017 Jan 30. EMBO J. 2017. PMID: 28137749 Free PMC article.
-
Munc18-bound syntaxin readily forms SNARE complexes with synaptobrevin in native plasma membranes.PLoS Biol. 2006 Oct;4(10):e330. doi: 10.1371/journal.pbio.0040330. PLoS Biol. 2006. PMID: 17002520 Free PMC article.
-
Neuronal SNARE complex assembly guided by Munc18-1 and Munc13-1.FEBS Open Bio. 2022 Nov;12(11):1939-1957. doi: 10.1002/2211-5463.13394. Epub 2022 Mar 22. FEBS Open Bio. 2022. PMID: 35278279 Free PMC article. Review.
-
Binding of Munc18-1 to synaptobrevin and to the SNARE four-helix bundle.Biochemistry. 2010 Mar 2;49(8):1568-76. doi: 10.1021/bi9021878. Biochemistry. 2010. PMID: 20102228 Free PMC article.
-
Molecular Mechanisms Underlying Neurotransmitter Release.Annu Rev Biophys. 2022 May 9;51:377-408. doi: 10.1146/annurev-biophys-111821-104732. Epub 2022 Feb 15. Annu Rev Biophys. 2022. PMID: 35167762 Free PMC article. Review.
References
-
- Abraham MJ, Murtola T, Schulz R, Páll S, Smith JC, Hess B, et al. GROMACS: high performance molecular simulations through multi‐level parallelism from laptops to supercomputers. SoftwareX. 2015;1–2:19–25.
-
- Anderson RJ, Weng Z, Campbell RK, Jiang X. Main‐chain conformational tendencies of amino acids. Proteins. 2005;60:679–689. - PubMed
-
- Archbold JK, Whitten AE, Hu S‐H, Collins BM, Martin JL. SNARE‐ing the structures of Sec1/Munc18 proteins. Curr Opin Struct Biol. 2014;29:44–51. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
