

A novel mucosal bivalent vaccine of EV-A71/EV-D68 adjuvanted with polysaccharides from Ganoderma lucidum protects mice against EV-A71 and EV-D68 lethal challenge
- PMID: 38110940
- PMCID: PMC10729491
- DOI: 10.1186/s12929-023-00987-3
A novel mucosal bivalent vaccine of EV-A71/EV-D68 adjuvanted with polysaccharides from Ganoderma lucidum protects mice against EV-A71 and EV-D68 lethal challenge
Abstract
Background: Human enteroviruses A71 (EV-A71) and D68 (EV-D68) are the suspected causative agents of hand-foot-and-mouth disease, aseptic meningitis, encephalitis, acute flaccid myelitis, and acute flaccid paralysis in children. Until now, no cure nor mucosal vaccine existed for EV-A71 and EV-D68. Novel mucosal bivalent vaccines are highly important for preventing EV-A71 and EV-D68 infections.
Methods: In this study, formalin-inactivated EV-A71 and EV-D68 were used as antigens, while PS-G, a polysaccharide from Ganoderma lucidum, was used as an adjuvant. Natural polysaccharides have the characteristics of intrinsic immunomodulation, biocompatibility, low toxicity, and safety. Mice were immunized intranasally with PBS, EV-A71, EV-D68, or EV-A71 + EV-D68, with or without PS-G as an adjuvant.
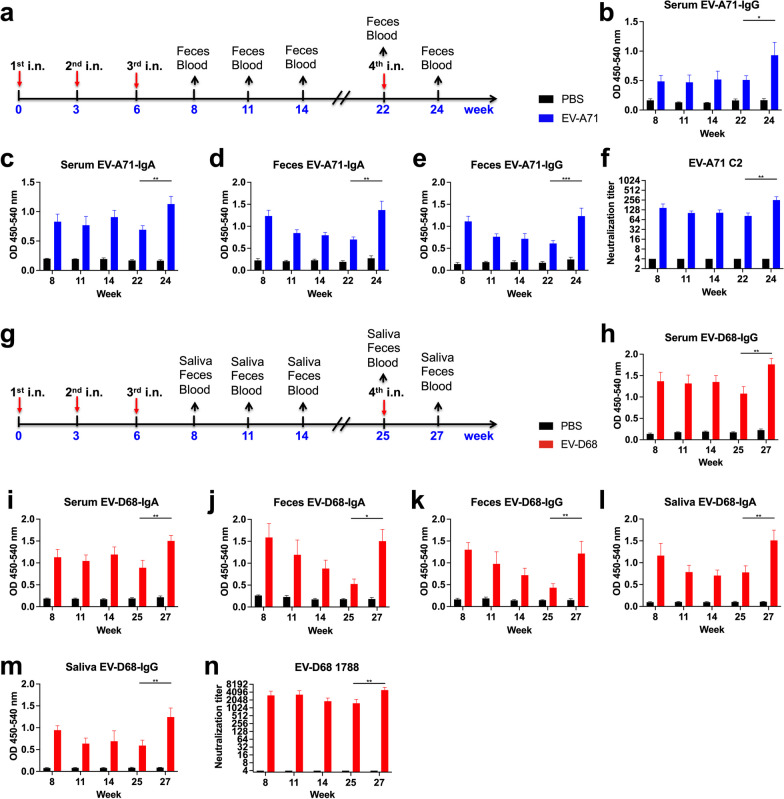
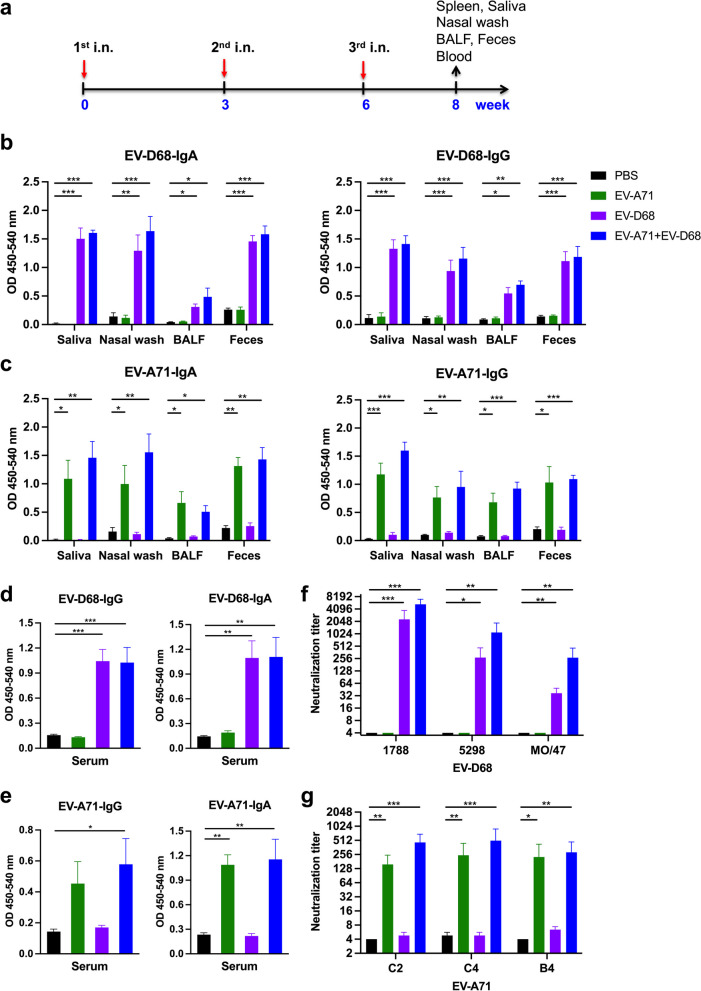
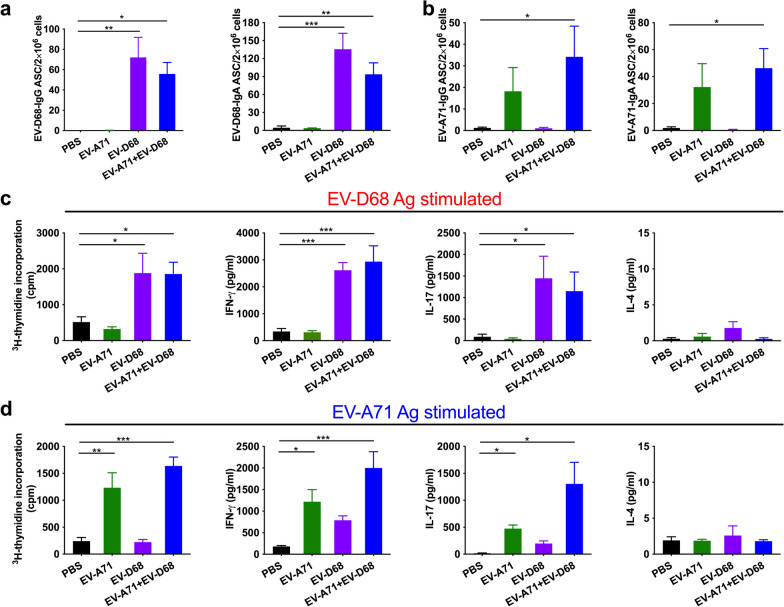
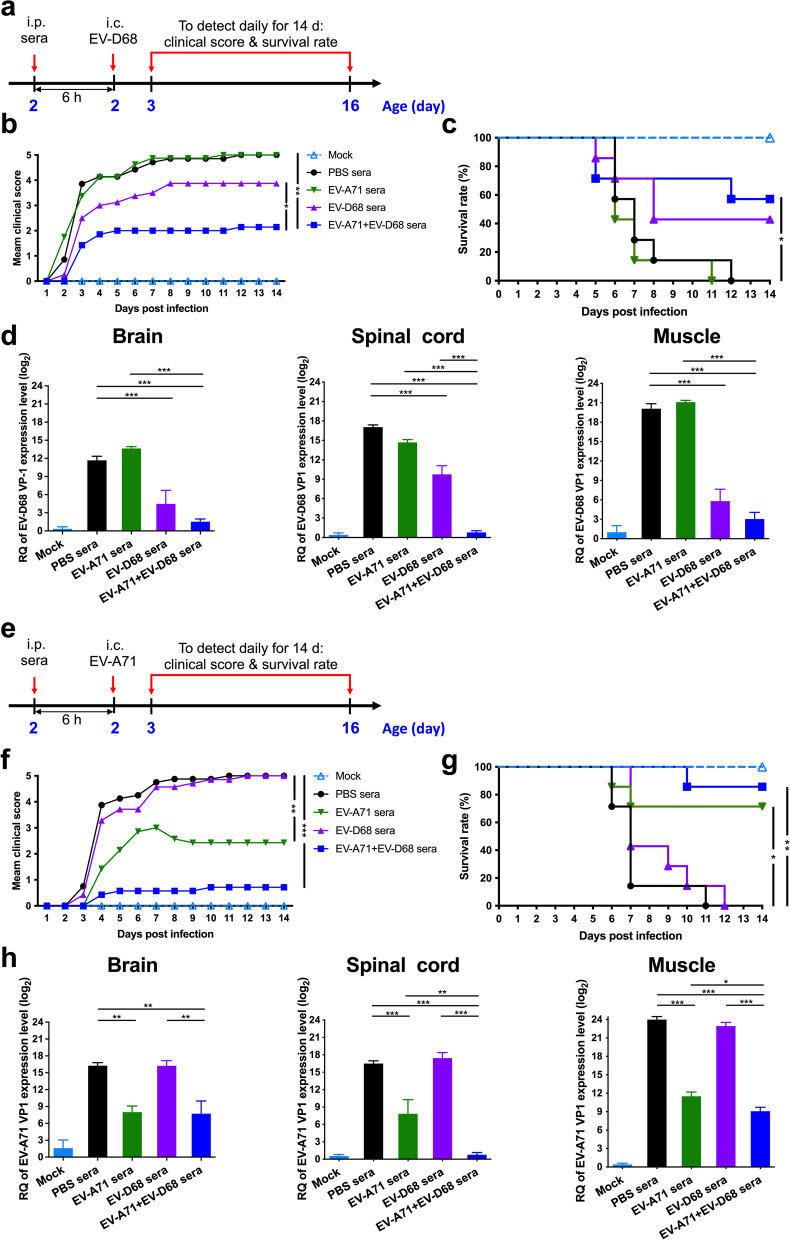
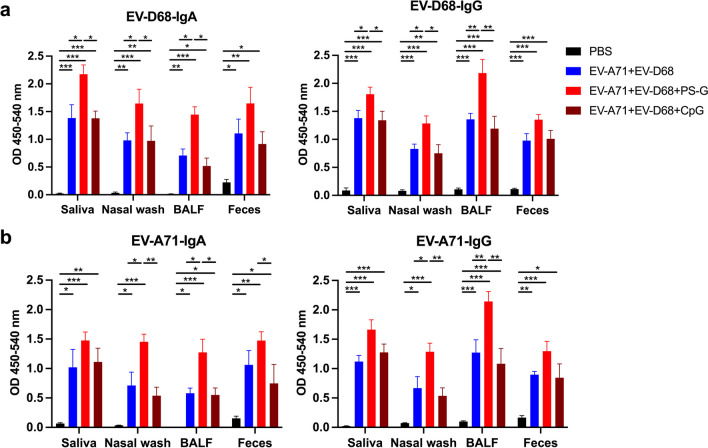
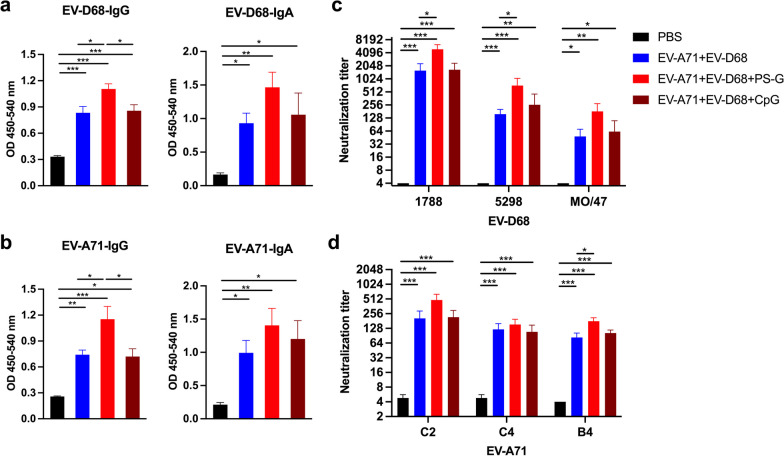
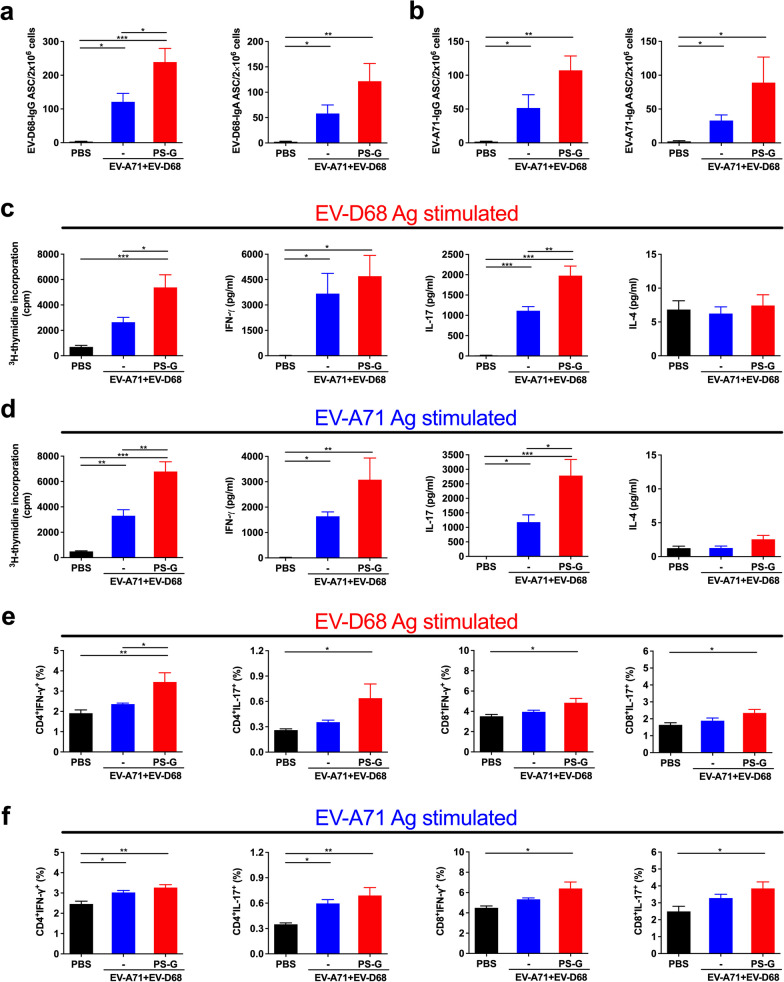
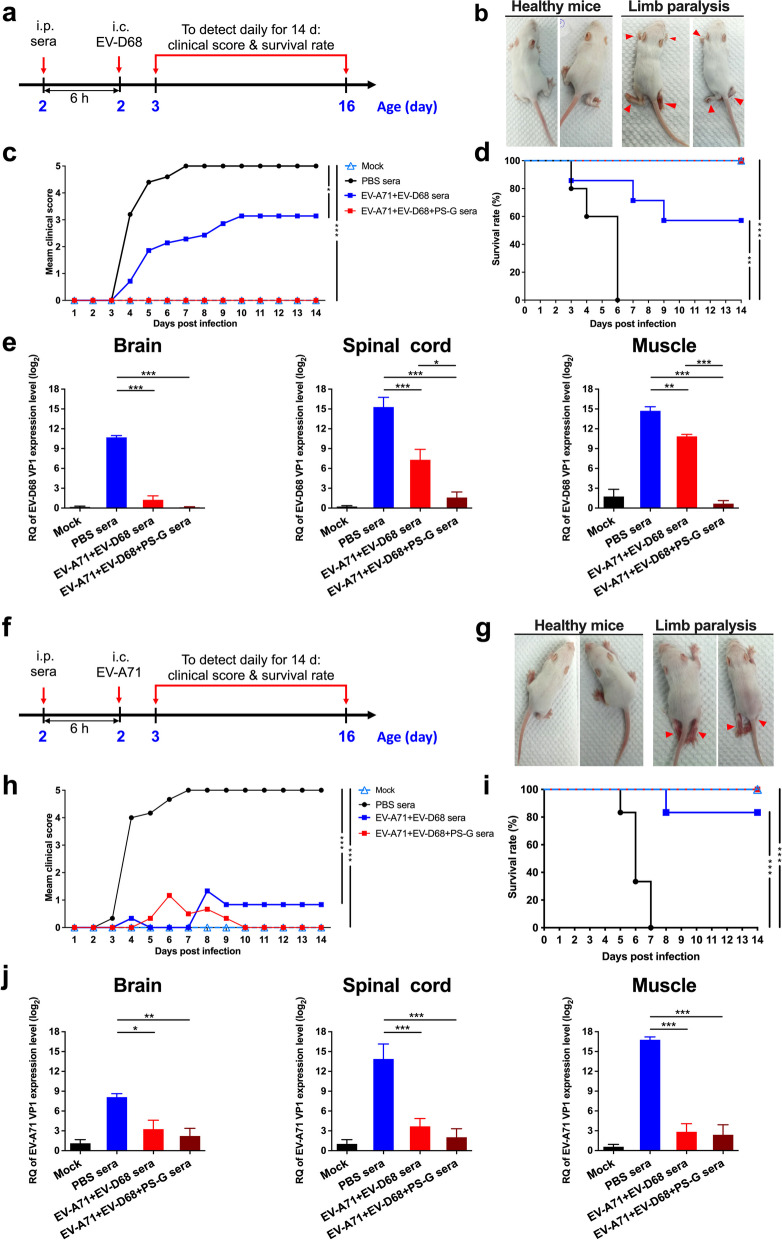
Results: The EV-A71 + EV-D68 bivalent vaccine generated considerable EV-A71- and EV-D68-specific IgG and IgA titres in the sera, nasal washes, saliva, bronchoalveolar lavage fluid, and feces. These antibodies neutralized EV-D68 and EV-A71 infectivity. They also cross-neutralized infections by different EV-D68 and EV-A71 sub-genotypes. Furthermore, compared with the PBS group, EV-A71 + EV-D68 + PS-G-vaccinated mice exhibited an increased number of EV-D68- and EV-A71-specific IgA- and IgG-producing cells. In addition, T-cell proliferative responses, and IFN-γ and IL-17 secretion in the spleen were substantially induced when PS-G was used as an adjuvant with EV-A71 + EV-D68. Finally, in vivo challenge experiments demonstrated that the immune sera induced by EV-A71 + EV-D68 + PS-G conferred protection in neonate mice against lethal EV-A71 and EV-D68 challenges as indicated by the increased survival rate and decreased clinical score and viral RNA tissue expression. Taken together, all EV-A71/EV-D68 + PS-G-immunized mice developed potent specific humoral, mucosal, and cellular immune responses to EV-D68 and EV-A71 and were protected against them.
Conclusions: These findings demonstrated that PS-G can be used as a potential adjuvant for EV-A71 and EV-D68 bivalent mucosal vaccines. Our results provide useful information for the further preclinical and clinical development of a mucosal bivalent enterovirus vaccine against both EV-A71 and EV-D68 infections.
Keywords: Acute flaccid myelitis; Acute flaccid paralysis; Adjuvant; Enterovirus A71; Enterovirus D68; Ganoderma lucidum polysaccharide; Intranasal; Mucosal vaccine.
© 2023. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interest.
Figures
Similar articles
-
A Polysaccharide Purified From Ganoderma lucidum Acts as a Potent Mucosal Adjuvant That Promotes Protective Immunity Against the Lethal Challenge With Enterovirus A71.Front Immunol. 2020 Sep 29;11:561758. doi: 10.3389/fimmu.2020.561758. eCollection 2020. Front Immunol. 2020. PMID: 33117346 Free PMC article.
-
Seroepidemiological and phylogenetic characterization of neurotropic enteroviruses in Ireland, 2005-2014.J Med Virol. 2017 Sep;89(9):1550-1558. doi: 10.1002/jmv.24765. Epub 2017 Jun 6. J Med Virol. 2017. PMID: 28071799
-
An inactivated whole-virion vaccine for Enterovirus D68 adjuvanted with CpG ODN or AddaVax elicits potent protective immunity in mice.Vaccine. 2024 Apr 2;42(9):2463-2474. doi: 10.1016/j.vaccine.2024.03.016. Epub 2024 Mar 11. Vaccine. 2024. PMID: 38472067
-
Regulation of the proteostasis network during enterovirus infection: A feedforward mechanism for EV-A71 and EV-D68.Antiviral Res. 2021 Apr;188:105019. doi: 10.1016/j.antiviral.2021.105019. Epub 2021 Jan 20. Antiviral Res. 2021. PMID: 33484748 Review.
-
Epidemiology of acute flaccid myelitis in children in the Netherlands, 2014 to 2019.Euro Surveill. 2022 Oct;27(42):2200157. doi: 10.2807/1560-7917.ES.2022.27.42.2200157. Euro Surveill. 2022. PMID: 36268734 Free PMC article. Review.
Cited by
-
EV-D68 cleaves LARP1 and PABPC1 by 3Cpro to redirect host mRNA translation machinery toward its genomic RNA.PLoS Pathog. 2025 Apr 28;21(4):e1013098. doi: 10.1371/journal.ppat.1013098. eCollection 2025 Apr. PLoS Pathog. 2025. PMID: 40294010 Free PMC article.
-
Advances in the Treatment of Enterovirus-D68 and Rhinovirus Respiratory Infections.Infect Dis Rep. 2025 Jun 1;17(3):61. doi: 10.3390/idr17030061. Infect Dis Rep. 2025. PMID: 40559192 Free PMC article. Review.
-
Insights Into Enterovirus D68 Immunology: Unraveling the Mysteries of Host-Pathogen Interactions.Immun Inflamm Dis. 2025 Feb;13(2):e70117. doi: 10.1002/iid3.70117. Immun Inflamm Dis. 2025. PMID: 39912556 Free PMC article. Review.
-
Enterovirus-D68 - A Reemerging Non-Polio Enterovirus that Causes Severe Respiratory and Neurological Disease in Children.Front Virol. 2024;4:1328457. doi: 10.3389/fviro.2024.1328457. Epub 2024 Feb 14. Front Virol. 2024. PMID: 39246649 Free PMC article.
-
Pterostilbene Exhibits Broad-Spectrum Antiviral Activity by Targeting the Enterovirus Capsid, Inactivating Viral Particles, Blocking Viral Binding, and Protecting Mice From Lethal EV-A71 Challenge.Phytother Res. 2025 Apr 16;39(6):2672-88. doi: 10.1002/ptr.8496. Online ahead of print. Phytother Res. 2025. PMID: 40235439 Free PMC article.
References
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
