

An atypical two-component system, AtcR/AtcK, simultaneously regulates the biosynthesis of multiple secondary metabolites in Streptomyces bingchenggensis
- PMID: 38112424
- PMCID: PMC10807435
- DOI: 10.1128/aem.01300-23
An atypical two-component system, AtcR/AtcK, simultaneously regulates the biosynthesis of multiple secondary metabolites in Streptomyces bingchenggensis
Abstract
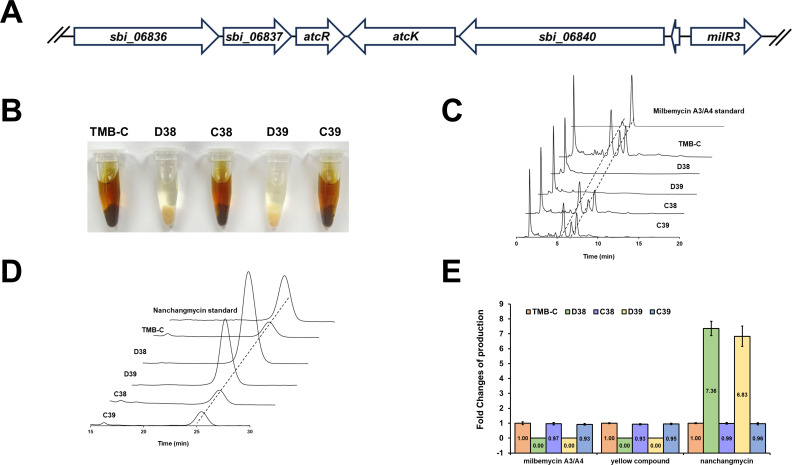
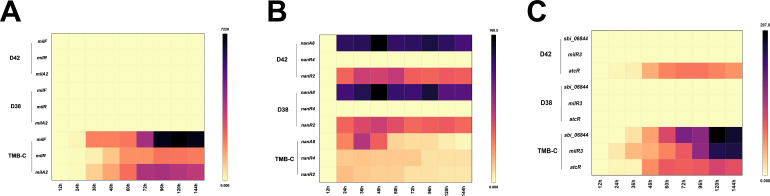
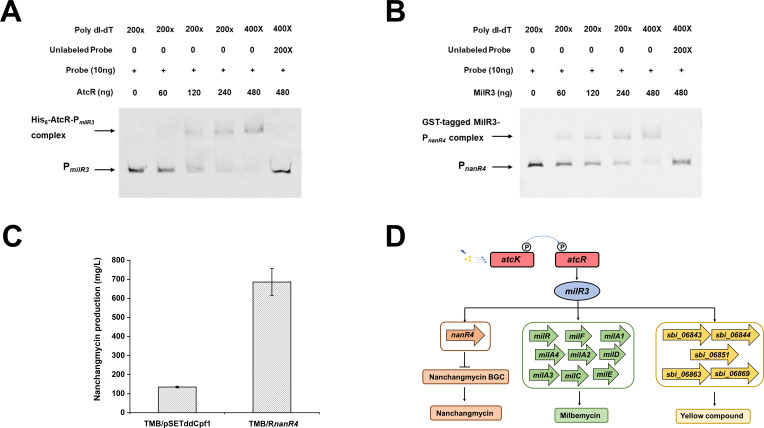
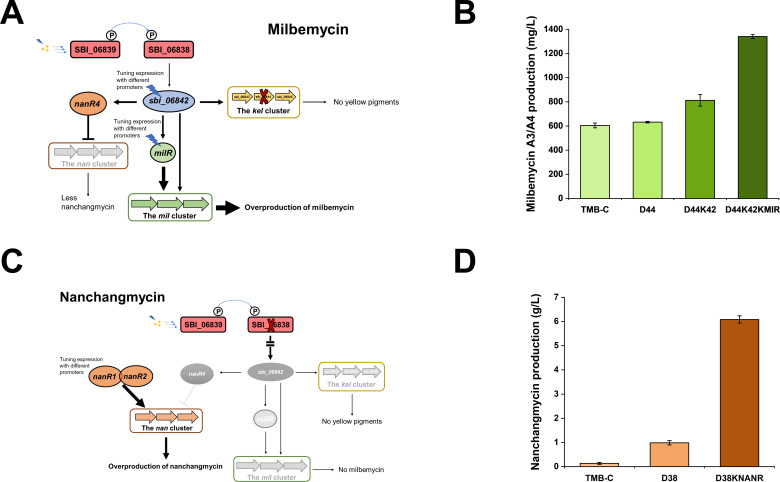
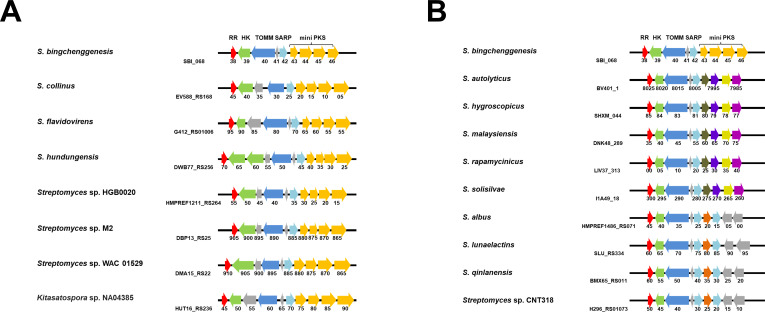
Streptomyces bingchenggensis is an industrial producer of milbemycins, which are important anthelmintic and insecticidal agents. Two-component systems (TCSs), which are typically situated in the same operon and are composed of a histidine kinase and a response regulator, are the predominant signal transduction pathways involved in the regulation of secondary metabolism in Streptomyces. Here, an atypical TCS, AtcR/AtcK, in which the encoding genes (sbi_06838/sbi_06839) are organized in a head-to-head pair, was demonstrated to be indispensable for the biosynthesis of multiple secondary metabolites in S. bingchenggensis. With the null TCS mutants, the production of milbemycin and yellow compound was abolished but nanchangmycin was overproduced. Transcriptional analysis and electrophoretic mobility shift assays showed that AtcR regulated the biosynthesis of these three secondary metabolites by a MilR3-mediated cascade. First, AtcR was activated by phosphorylation from signal-triggered AtcK. Second, the activated AtcR promoted the transcription of milR3. Third, MilR3 specifically activated the transcription of downstream genes from milbemycin and yellow compound biosynthetic gene clusters (BGCs) and nanR4 from the nanchangmycin BGC. Finally, because NanR4 is a specific repressor in the nanchangmycin BGC, activation of MilR3 downstream genes led to the production of yellow compound and milbemycin but inhibited nanchangmycin production. By rewiring the regulatory cascade, two strains were obtained, the yield of nanchangmycin was improved by 45-fold to 6.08 g/L and the production of milbemycin was increased twofold to 1.34 g/L. This work has broadened our knowledge on atypical TCSs and provided practical strategies to engineer strains for the production of secondary metabolites in Streptomyces.IMPORTANCEStreptomyces bingchenggensis is an important industrial strain that produces milbemycins. Two-component systems (TCSs), which consist of a histidine kinase and a response regulator, are the predominant signal transduction pathways involved in the regulation of secondary metabolism in Streptomyces. Coupled encoding genes of TCSs are typically situated in the same operon. Here, TCSs with encoding genes situated in separate head-to-head neighbor operons were labeled atypical TCSs. It was found that the atypical TCS AtcR/AtcK played an indispensable role in the biosynthesis of milbemycin, yellow compound, and nanchangmycin in S. bingchenggensis. This atypical TCS regulated the biosynthesis of specialized metabolites in a cascade mediated via a cluster-situated regulator, MilR3. Through rewiring the regulatory pathways, strains were successfully engineered to overproduce milbemycin and nanchangmycin. To the best of our knowledge, this is the first report on atypical TCS, in which the encoding genes of RR and HK were situated in separate head-to-head neighbor operons, involved in secondary metabolism. In addition, data mining showed that atypical TCSs were widely distributed in actinobacteria.
Keywords: Streptomyces bingchenggensis; milbemycins; nanchangmycin; rewiring regulatory network; strain engineering; two-component system.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
SbhR, a DeoR family regulator, modulates secondary metabolism via the atypical two-component system AtcK/R in Streptomyces bingchenggensis.Int J Biol Macromol. 2025 May;310(Pt 1):143161. doi: 10.1016/j.ijbiomac.2025.143161. Epub 2025 Apr 14. Int J Biol Macromol. 2025. PMID: 40239784
-
MilR3, a unique SARP family pleiotropic regulator in Streptomyces bingchenggensis.Arch Microbiol. 2022 Sep 19;204(10):631. doi: 10.1007/s00203-022-03240-x. Arch Microbiol. 2022. PMID: 36121479
-
Genetic engineering of Streptomyces bingchenggensis to produce milbemycins A3/A4 as main components and eliminate the biosynthesis of nanchangmycin.Appl Microbiol Biotechnol. 2013 Dec;97(23):10091-101. doi: 10.1007/s00253-013-5255-5. Epub 2013 Sep 29. Appl Microbiol Biotechnol. 2013. PMID: 24077727
-
Recent advances in the research of milbemycin biosynthesis and regulation as well as strategies for strain improvement.Arch Microbiol. 2021 Dec;203(10):5849-5857. doi: 10.1007/s00203-021-02575-1. Epub 2021 Sep 22. Arch Microbiol. 2021. PMID: 34550409 Review.
-
Trends in the two-component system's role in the synthesis of antibiotics by Streptomyces.Appl Microbiol Biotechnol. 2023 Aug;107(15):4727-4743. doi: 10.1007/s00253-023-12623-z. Epub 2023 Jun 21. Appl Microbiol Biotechnol. 2023. PMID: 37341754 Free PMC article. Review.
Cited by
-
Mechanisms and Impact of Rhizosphere Microbial Metabolites on Crop Health, Traits, Functional Components: A Comprehensive Review.Molecules. 2024 Dec 15;29(24):5922. doi: 10.3390/molecules29245922. Molecules. 2024. PMID: 39770010 Free PMC article. Review.
-
The roles of SARP family regulators involved in secondary metabolism in Streptomyces.Front Microbiol. 2024 Mar 14;15:1368809. doi: 10.3389/fmicb.2024.1368809. eCollection 2024. Front Microbiol. 2024. PMID: 38550856 Free PMC article. Review.
-
A Rho-Type GTPase Protein Pocdc42 Regulates the Development and Cellulase Expression of Penicillium oxalicum.Curr Microbiol. 2025 Jul 6;82(8):365. doi: 10.1007/s00284-025-04355-2. Curr Microbiol. 2025. PMID: 40617885
-
Overexpression of the Wbl family regulator whiDsbh modulates secondary metabolite biosynthesis in Streptomyces.World J Microbiol Biotechnol. 2025 May 15;41(5):176. doi: 10.1007/s11274-025-04400-z. World J Microbiol Biotechnol. 2025. PMID: 40369383
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- LY19C010002/MOST | NSFC | NSFC-Zhejiang Joint Fund | | Natural Science Foundation of Zhejiang Province (ZJNSF)
- 2001xg07/the Scientific Research Foundation of Taizhou
- 21nya20/the Scientific Research Foundation of Taizhou
- 22nya12/the Scientific Research Foundation of Taizhou
- 23gya07/the Scientific Research Foundation of Taizhou
LinkOut - more resources
Full Text Sources
