

Molecular traces of Drosophila hemocytes reveal transcriptomic conservation with vertebrate myeloid cells
- PMID: 38113249
- PMCID: PMC10763942
- DOI: 10.1371/journal.pgen.1011077
Molecular traces of Drosophila hemocytes reveal transcriptomic conservation with vertebrate myeloid cells
Abstract
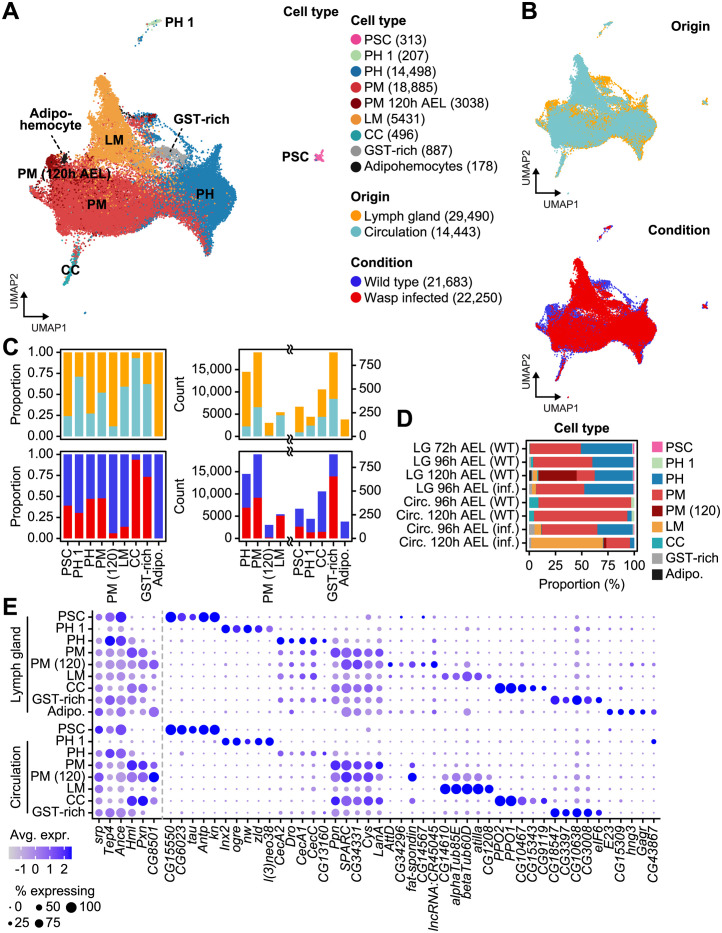
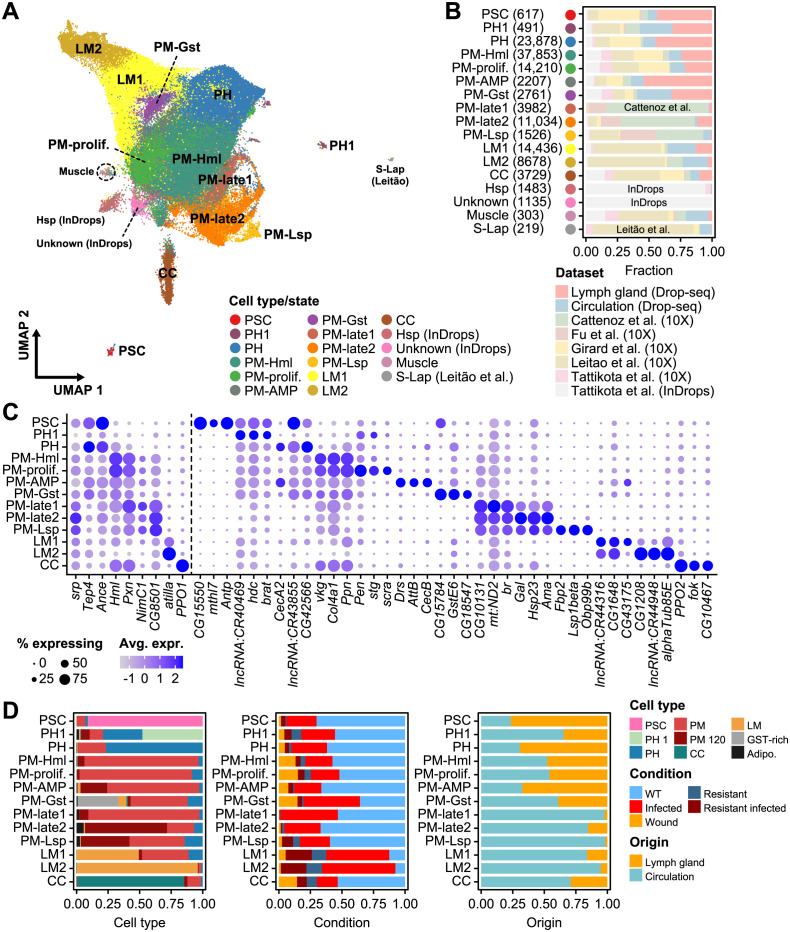
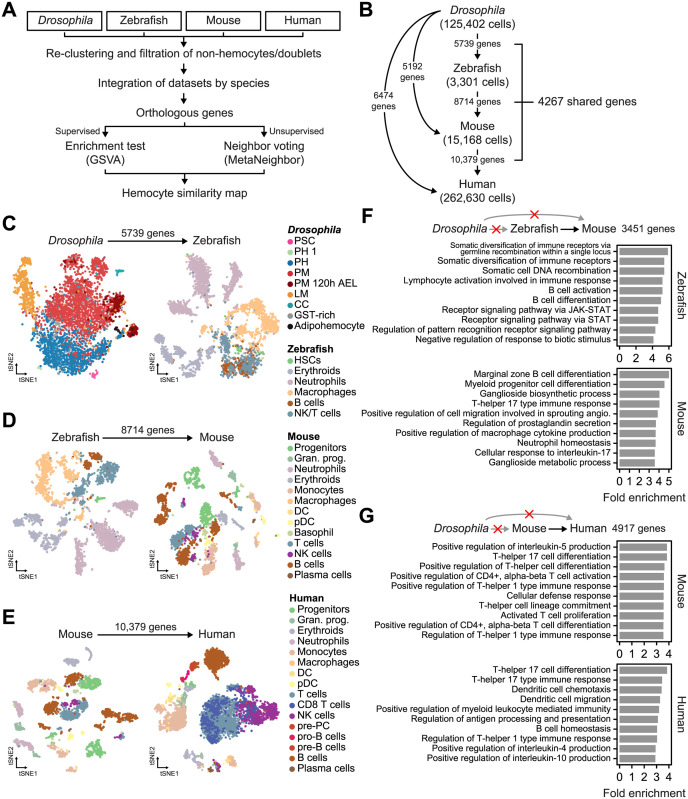
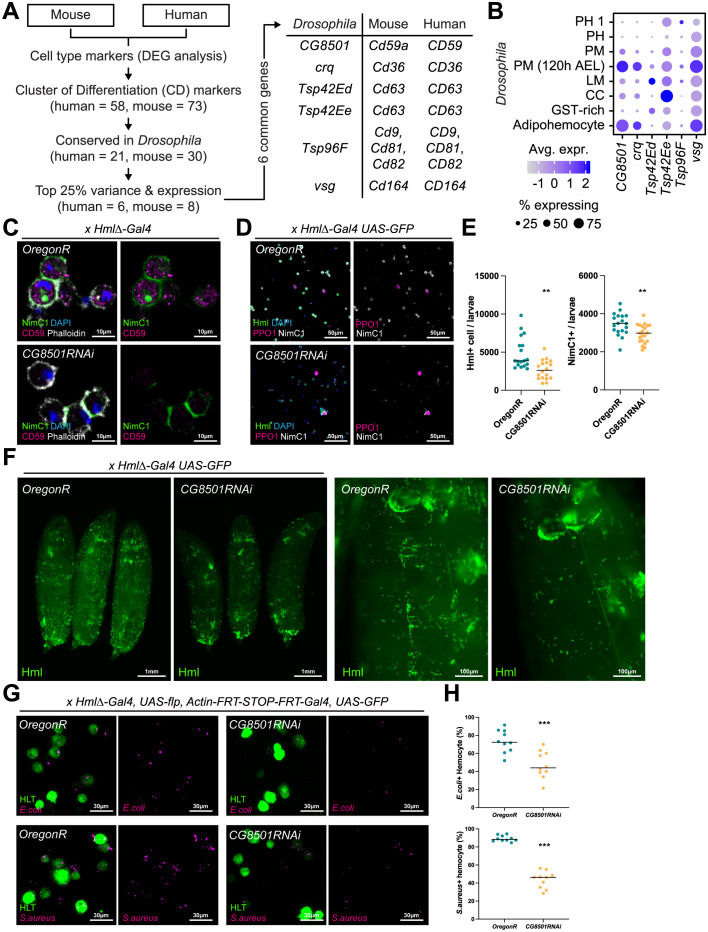
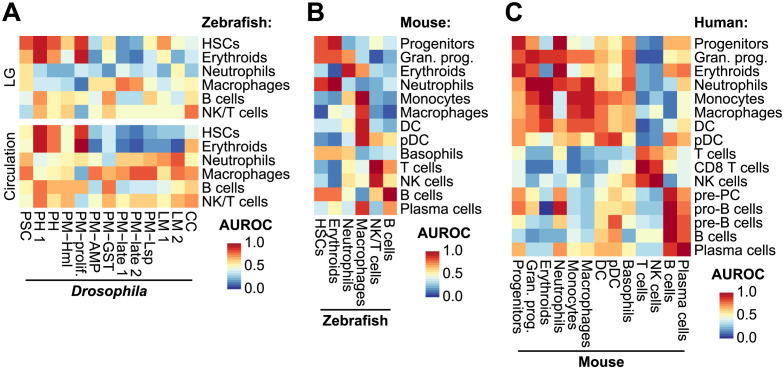
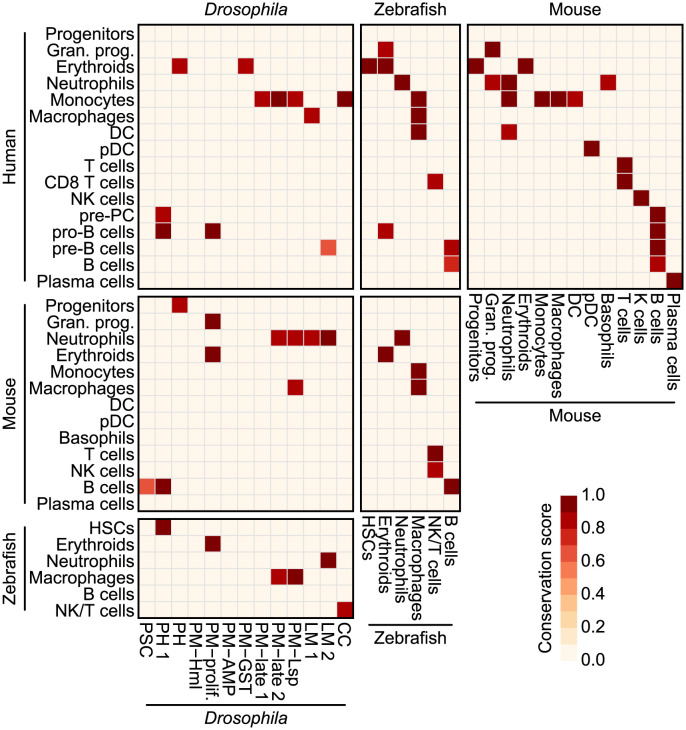
Drosophila hemocytes serve as the primary defense system against harmful threats, allowing the animals to thrive. Hemocytes are often compared to vertebrate innate immune system cells due to the observed functional similarities between the two. However, the similarities have primarily been established based on a limited number of genes and their functional homologies. Thus, a systematic analysis using transcriptomic data could offer novel insights into Drosophila hemocyte function and provide new perspectives on the evolution of the immune system. Here, we performed cross-species comparative analyses using single-cell RNA sequencing data from Drosophila and vertebrate immune cells. We found several conserved markers for the cluster of differentiation (CD) genes in Drosophila hemocytes and validated the role of CG8501 (CD59) in phagocytosis by plasmatocytes, which function much like macrophages in vertebrates. By comparing whole transcriptome profiles in both supervised and unsupervised analyses, we showed that Drosophila hemocytes are largely homologous to vertebrate myeloid cells, especially plasmatocytes to monocytes/macrophages and prohemocyte 1 (PH1) to hematopoietic stem cells. Furthermore, a small subset of prohemocytes with hematopoietic potential displayed homology with hematopoietic progenitor populations in vertebrates. Overall, our results provide a deeper understanding of molecular conservation in the Drosophila immune system.
Copyright: © 2023 Yoon et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Single-cell transcriptome maps of myeloid blood cell lineages in Drosophila.Nat Commun. 2020 Sep 8;11(1):4483. doi: 10.1038/s41467-020-18135-y. Nat Commun. 2020. PMID: 32900993 Free PMC article.
-
Hemocyte Nuclei Isolation from Adult Drosophila melanogaster for snRNA-seq.Methods Mol Biol. 2024;2713:71-79. doi: 10.1007/978-1-0716-3437-0_4. Methods Mol Biol. 2024. PMID: 37639115
-
Distinctive features of Zaprionus indianus hemocyte differentiation and function revealed by transcriptomic analysis.Front Immunol. 2023 Dec 21;14:1322381. doi: 10.3389/fimmu.2023.1322381. eCollection 2023. Front Immunol. 2023. PMID: 38187383 Free PMC article.
-
Hematopoietic plasticity mapped in Drosophila and other insects.Elife. 2022 Aug 3;11:e78906. doi: 10.7554/eLife.78906. Elife. 2022. PMID: 35920811 Free PMC article. Review.
-
Drosophila as a Model to Study Cellular Communication Between the Hematopoietic Niche and Blood Progenitors Under Homeostatic Conditions and in Response to an Immune Stress.Front Immunol. 2021 Aug 16;12:719349. doi: 10.3389/fimmu.2021.719349. eCollection 2021. Front Immunol. 2021. PMID: 34484226 Free PMC article. Review.
Cited by
-
Editorial: Community series in recent advances in Drosophila cellular and humoral innate immunity: volume II.Front Immunol. 2024 Jun 18;15:1416296. doi: 10.3389/fimmu.2024.1416296. eCollection 2024. Front Immunol. 2024. PMID: 38957463 Free PMC article. No abstract available.
-
The NF-κB Factor Relish maintains blood progenitor homeostasis in the developing Drosophila lymph gland.PLoS Genet. 2024 Sep 9;20(9):e1011403. doi: 10.1371/journal.pgen.1011403. eCollection 2024 Sep. PLoS Genet. 2024. PMID: 39250509 Free PMC article.
-
Evolution of innate immunity: lessons from mammalian models shaping our current view of insect immunity.J Comp Physiol B. 2024 Apr;194(2):105-119. doi: 10.1007/s00360-024-01549-1. Epub 2024 Apr 4. J Comp Physiol B. 2024. PMID: 38573502 Review.
-
Environmental Diagnosis through a Flow Cytometric Approach.Int J Mol Sci. 2024 Oct 15;25(20):11069. doi: 10.3390/ijms252011069. Int J Mol Sci. 2024. PMID: 39456852 Free PMC article. Review.
-
Drosophila immune cells transport oxygen through PPO2 protein phase transition.Nature. 2024 Jul;631(8020):350-359. doi: 10.1038/s41586-024-07583-x. Epub 2024 Jun 26. Nature. 2024. PMID: 38926577 Free PMC article.
References
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
