

NET formation is a default epigenetic program controlled by PAD4 in apoptotic neutrophils
- PMID: 38117877
- PMCID: PMC10732518
- DOI: 10.1126/sciadv.adj1397
NET formation is a default epigenetic program controlled by PAD4 in apoptotic neutrophils
Abstract
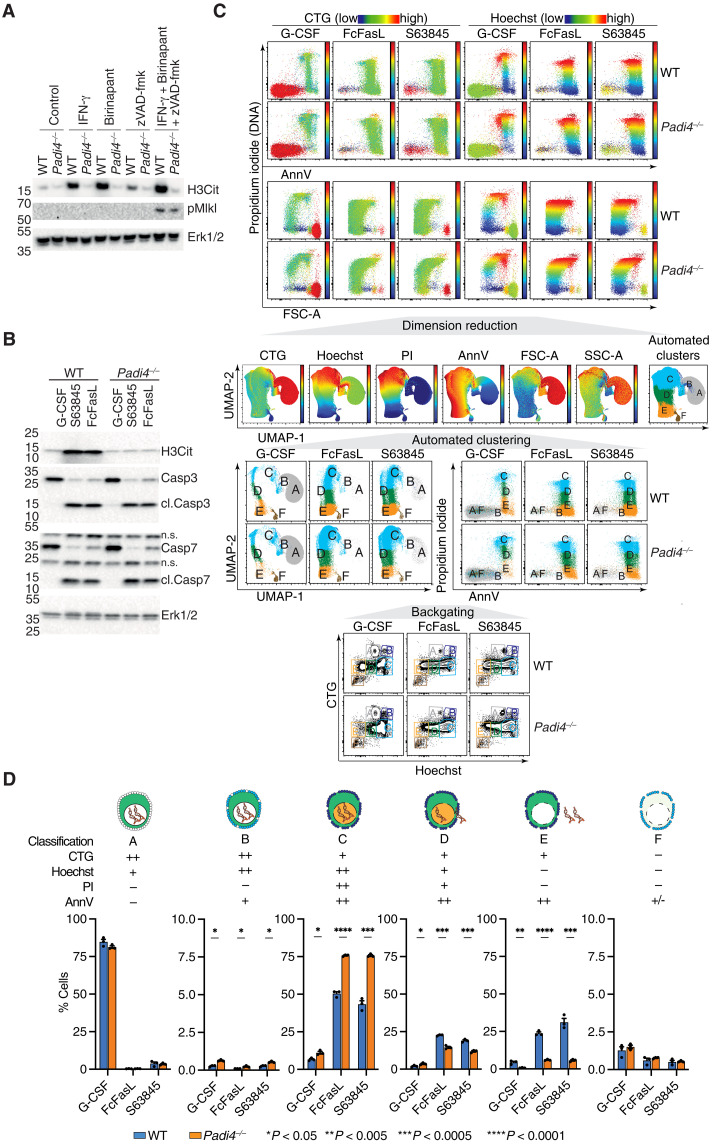
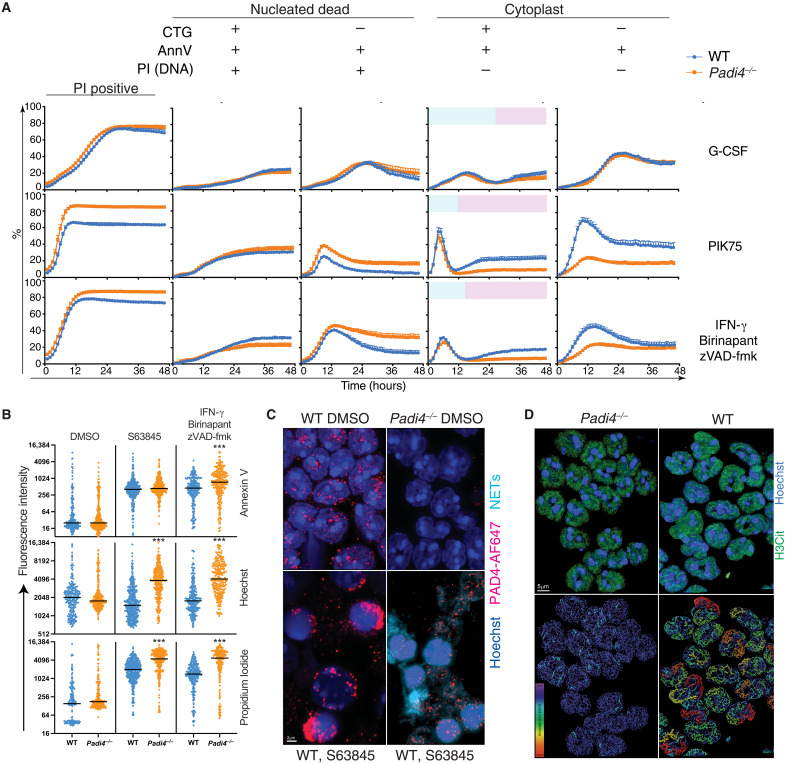
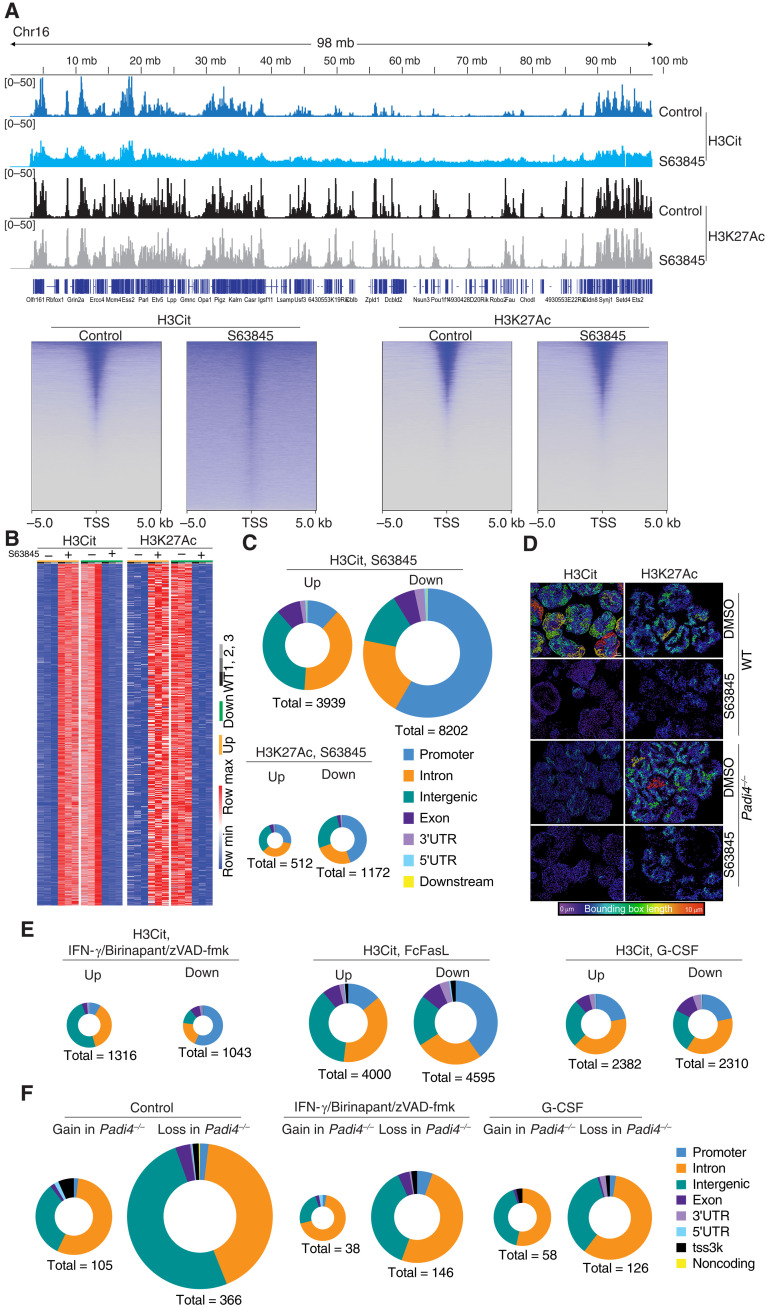
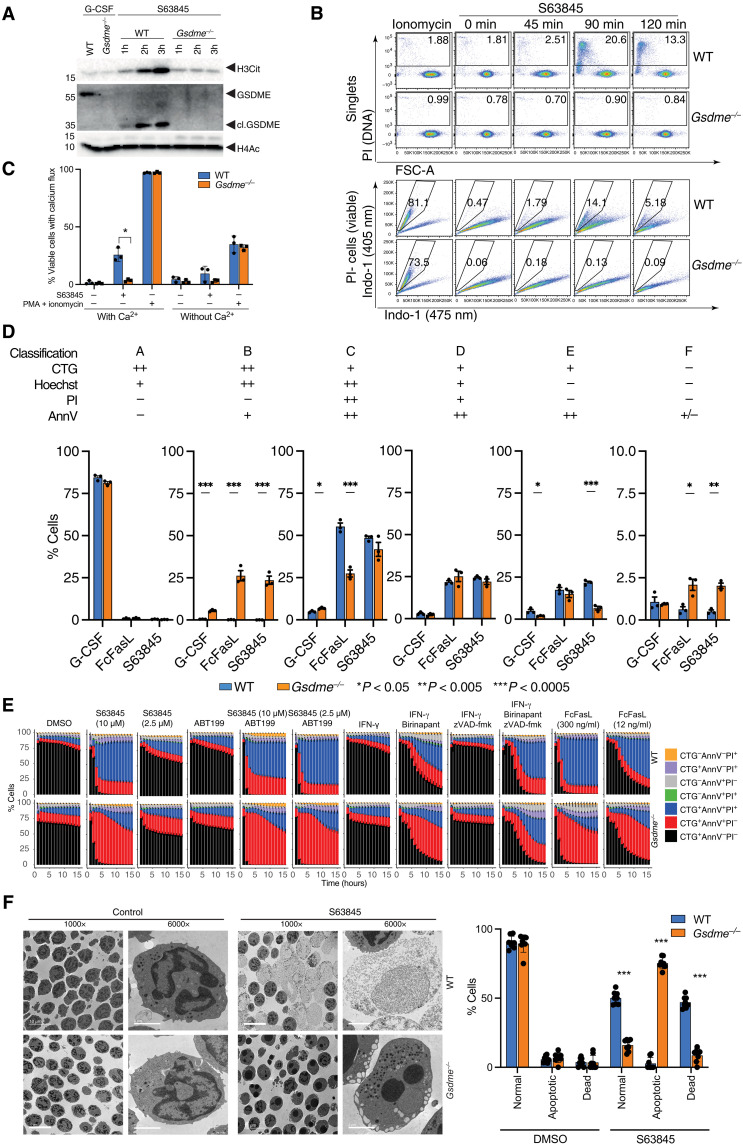
Neutrophil extracellular traps (NETs) not only counteract bacterial and fungal pathogens but can also promote thrombosis, autoimmunity, and sterile inflammation. The presence of citrullinated histones, generated by the peptidylarginine deiminase 4 (PAD4), is synonymous with NETosis and is considered independent of apoptosis. Mitochondrial- and death receptor-mediated apoptosis promote gasdermin E (GSDME)-dependent calcium mobilization and membrane permeabilization leading to histone H3 citrullination (H3Cit), nuclear DNA extrusion, and cytoplast formation. H3Cit is concentrated at the promoter in bone marrow neutrophils and redistributes in a coordinated process from promoter to intergenic and intronic regions during apoptosis. Loss of GSDME prevents nuclear and plasma membrane disruption of apoptotic neutrophils but prolongs early apoptosis-induced cellular changes to the chromatin and cytoplasmic granules. Apoptotic signaling engages PAD4 in neutrophils, establishing a cellular state that is primed for NETosis, but that occurs only upon membrane disruption by GSDME, thereby redefining the end of life for neutrophils.
Figures
References
-
- D’Cruz A. A., Speir M., Bliss-Moreau M., Dietrich S., Wang S., Chen A. A., Gavillet M., Al-Obeidi A., Lawlor K. E., Vince J. E., Kelliher M. A., Hakem R., Pasparakis M., Williams D. A., Ericsson M., Croker B. A., The pseudokinase MLKL activates PAD4-dependent NET formation in necroptotic neutrophils. Sci. Signal. 11, eaao1716 (2018). - PMC - PubMed
-
- Chen K. W., Monteleone M., Boucher D., Sollberger G., Ramnath D., Condon N. D., von Pein J. B., Broz P., Sweet M. J., Schroder K., Noncanonical inflammasome signaling elicits gasdermin D-dependent neutrophil extracellular traps. Sci. Immunol. 3, (2018). - PubMed
-
- Sollberger G., Choidas A., Burn G. L., Habenberger P., Di Lucrezia R., Kordes S., Menninger S., Eickhoff J., Nussbaumer P., Klebl B., Kruger R., Herzig A., Zychlinsky A., Gasdermin D plays a vital role in the generation of neutrophil extracellular traps. Sci. Immunol. 3, (2018). - PubMed
-
- Chen X., Shen Y., Draper W., Buenrostro J. D., Litzenburger U., Cho S. W., Satpathy A. T., Carter A. C., Ghosh R. P., East-Seletsky A., Doudna J. A., Greenleaf W. J., Liphardt J. T., Chang H. Y., ATAC-see reveals the accessible genome by transposase-mediated imaging and sequencing. Nat. Methods 13, 1013–1020 (2016). - PMC - PubMed
