

Reassessing endothelial-to-mesenchymal transition in mouse bone marrow: insights from lineage tracing models
- PMID: 38123537
- PMCID: PMC10733381
- DOI: 10.1038/s41467-023-44312-w
Reassessing endothelial-to-mesenchymal transition in mouse bone marrow: insights from lineage tracing models
Abstract
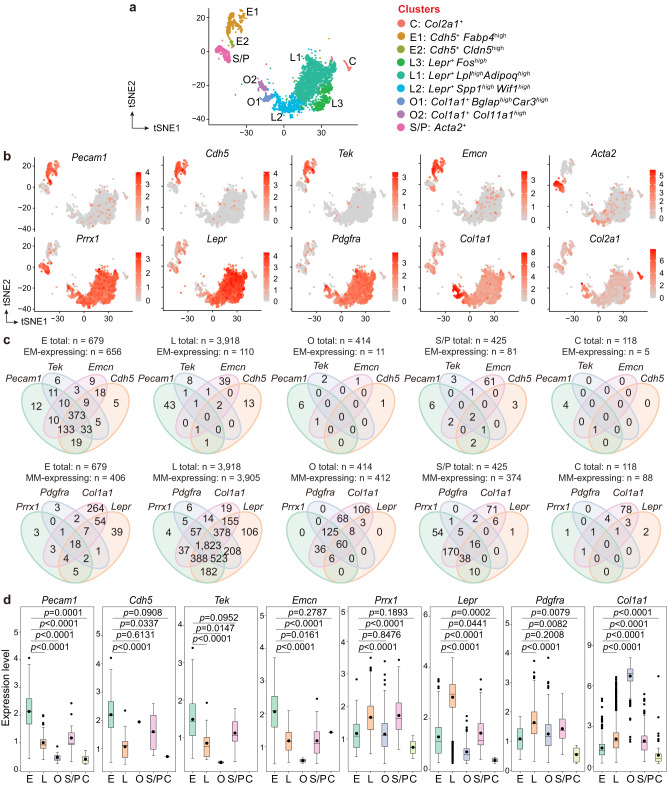
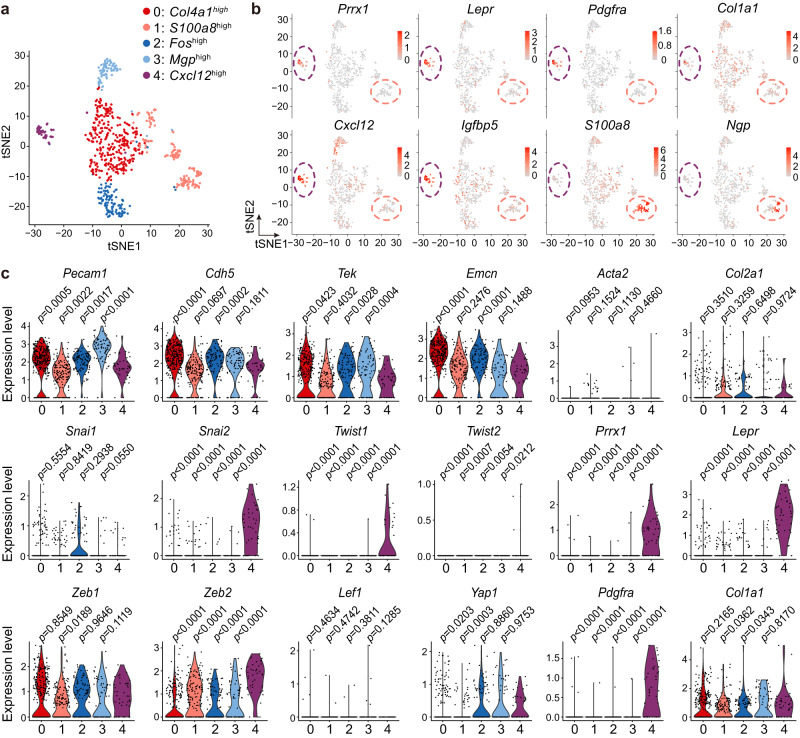
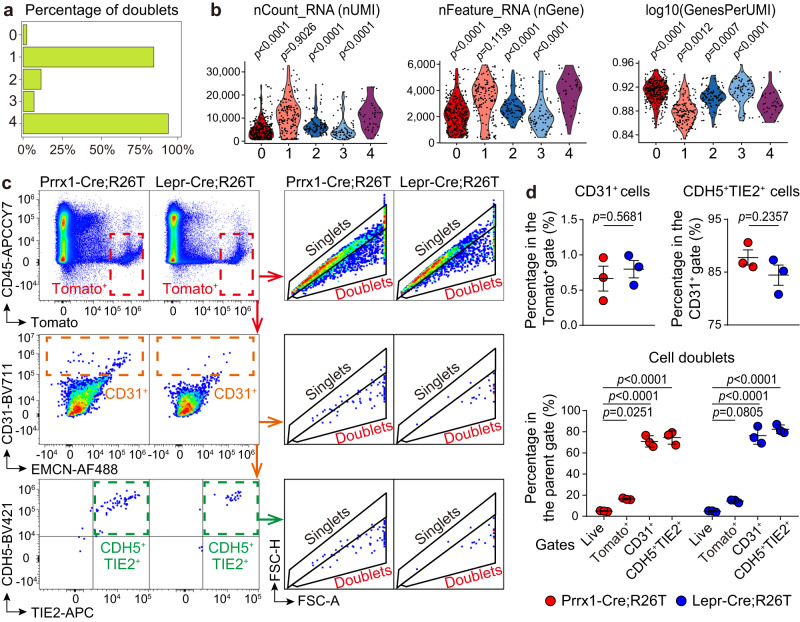
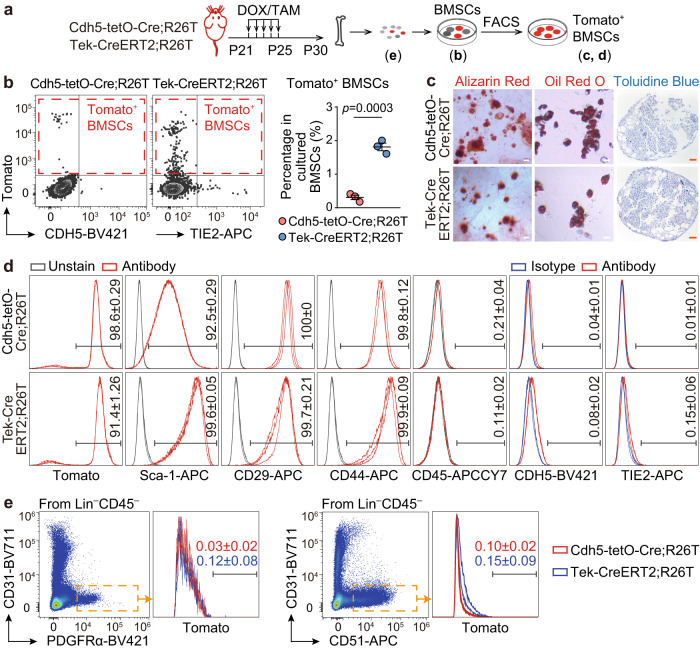
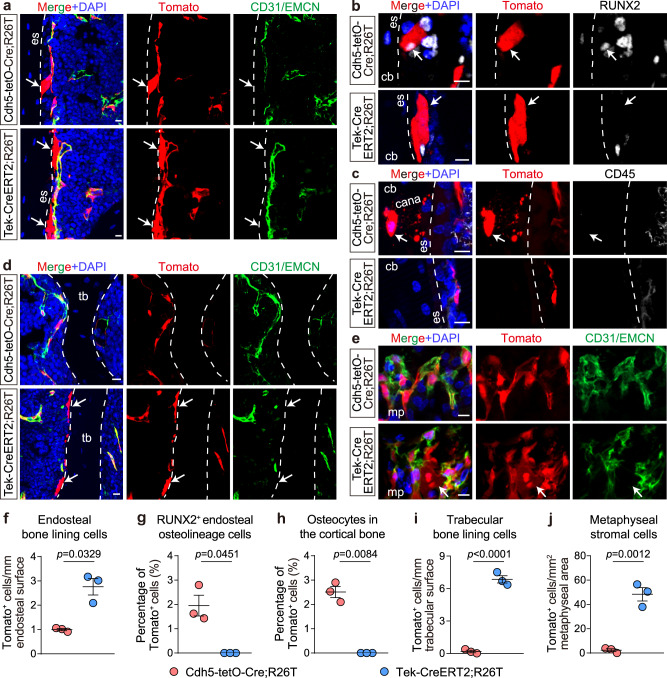
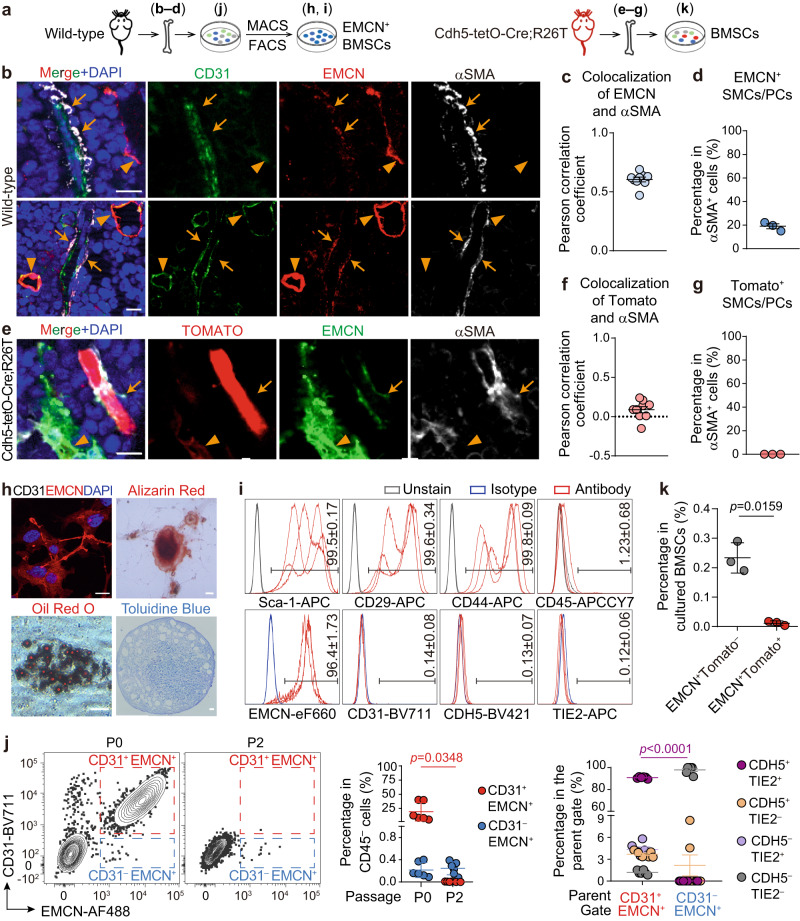
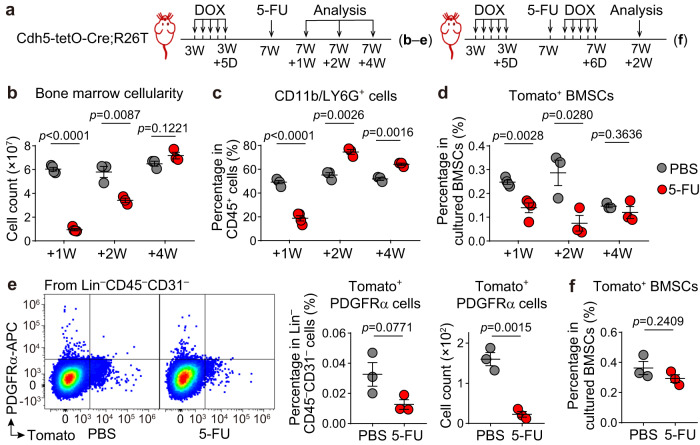
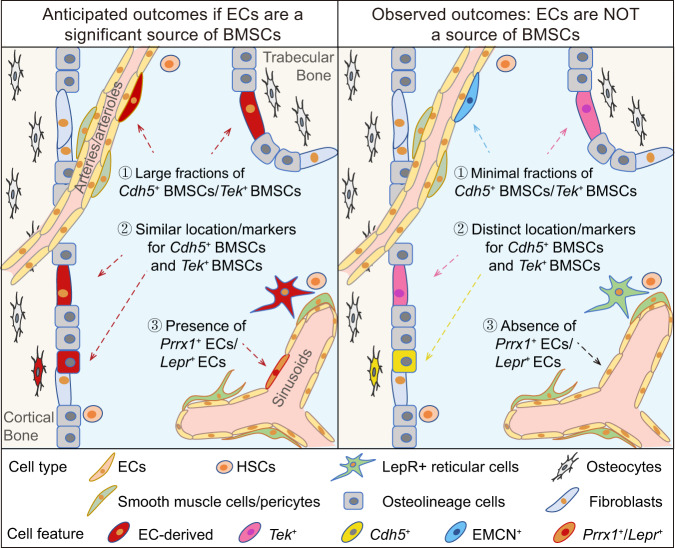
Endothelial cells (ECs) and bone marrow stromal cells (BMSCs) play crucial roles in supporting hematopoiesis and hematopoietic regeneration. However, whether ECs are a source of BMSCs remains unclear. Here, we evaluate the contribution of endothelial-to-mesenchymal transition to BMSC generation in postnatal mice. Single-cell RNA sequencing identifies ECs expressing BMSC markers Prrx1 and Lepr; however, this could not be validated using Prrx1-Cre and Lepr-Cre transgenic mice. Additionally, only a minority of BMSCs are marked by EC lineage tracing models using Cdh5-rtTA-tetO-Cre or Tek-CreERT2. Moreover, Cdh5+ BMSCs and Tek+ BMSCs show distinct spatial distributions and characteristic mesenchymal markers, suggestive of their origination from different progenitors rather than CDH5+ TEK+ ECs. Furthermore, myeloablation induced by 5-fluorouracil treatment does not increase Cdh5+ BMSCs. Our findings indicate that ECs hardly convert to BMSCs during homeostasis and myeloablation-induced hematopoietic regeneration, highlighting the importance of using appropriate genetic models and conducting careful data interpretation in studies concerning endothelial-to-mesenchymal transition.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
- 82072504/National Natural Science Foundation of China (National Science Foundation of China)
- 82125023/National Natural Science Foundation of China (National Science Foundation of China)
- 81600699/National Natural Science Foundation of China (National Science Foundation of China)
- 82272562/National Natural Science Foundation of China (National Science Foundation of China)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
