

Autophagic dysfunction and gut microbiota dysbiosis cause chronic immune activation in a Drosophila model of Gaucher disease
- PMID: 38127816
- PMCID: PMC10734978
- DOI: 10.1371/journal.pgen.1011063
Autophagic dysfunction and gut microbiota dysbiosis cause chronic immune activation in a Drosophila model of Gaucher disease
Abstract
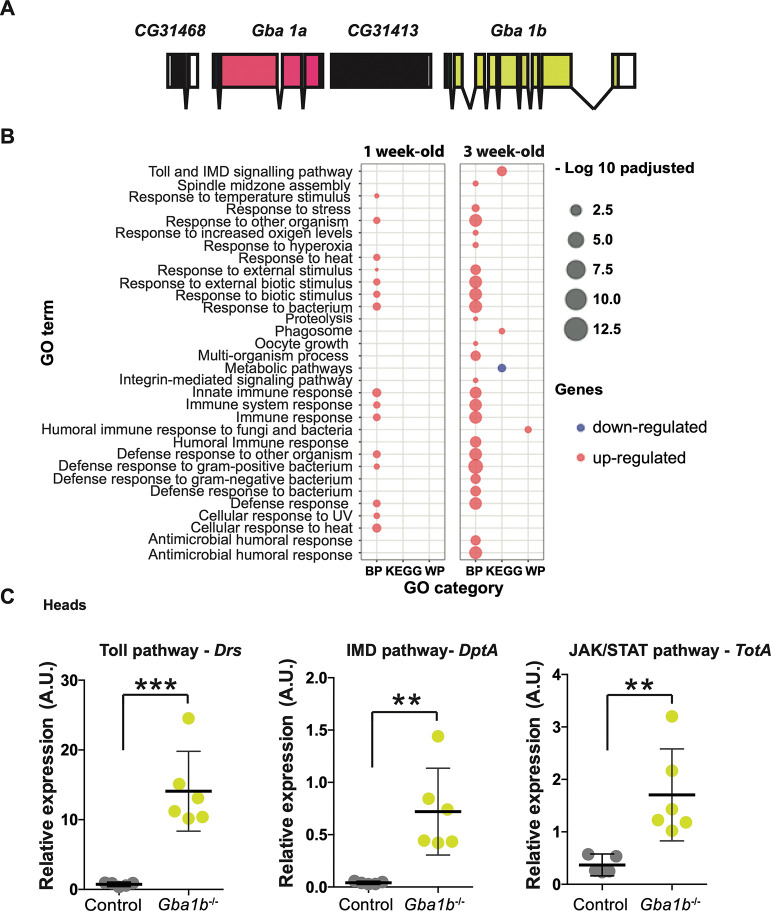
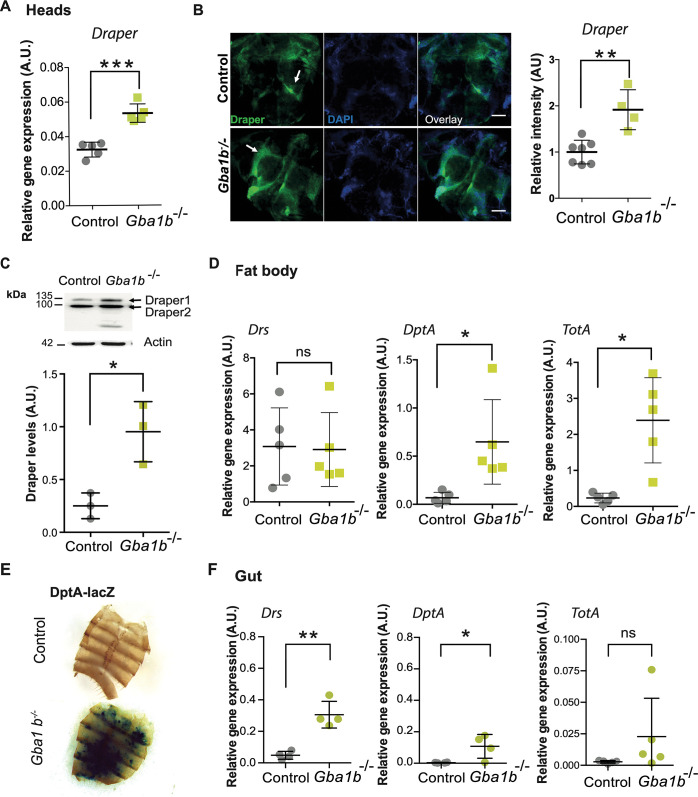
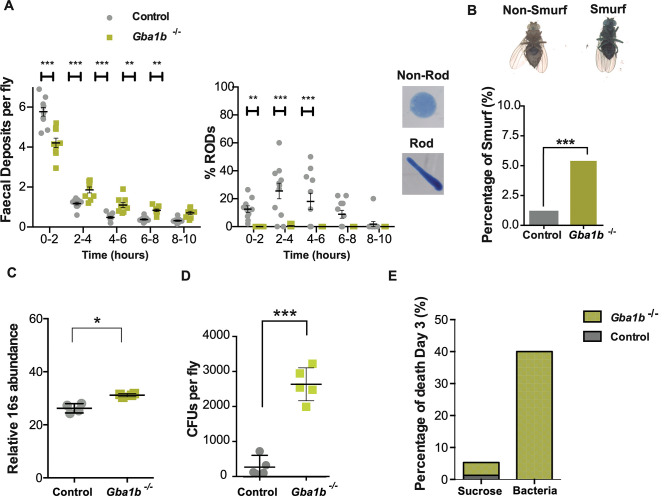
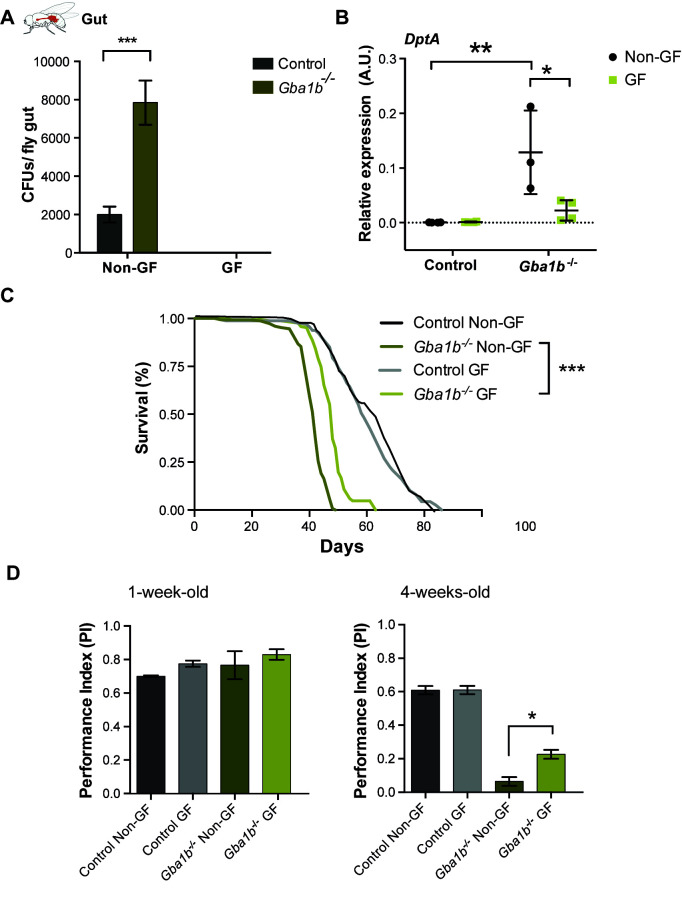
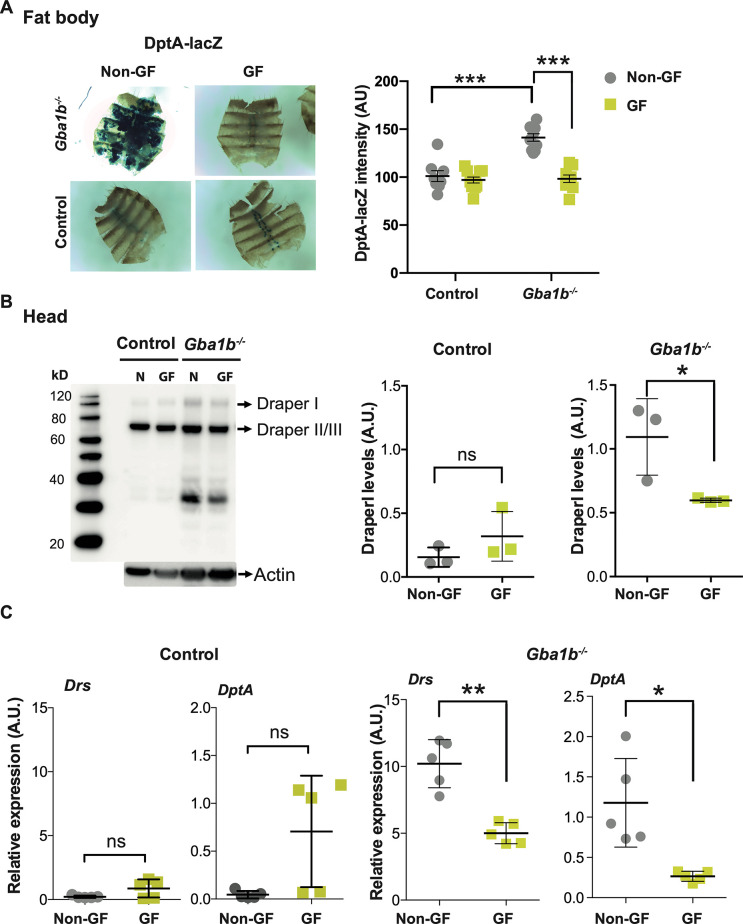
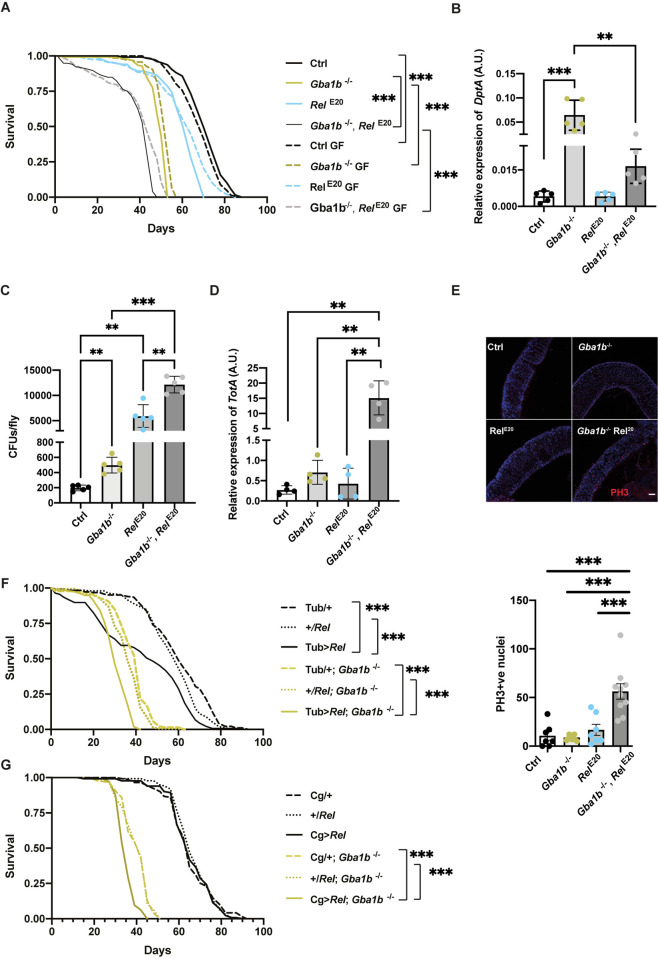
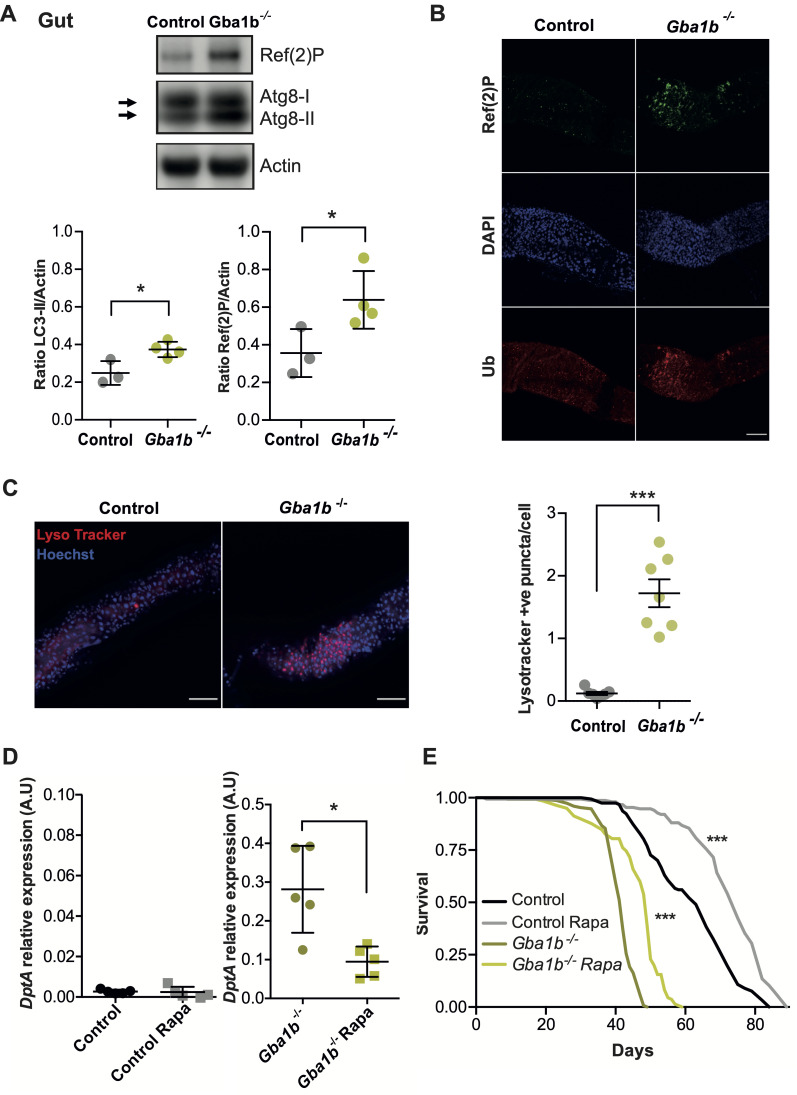
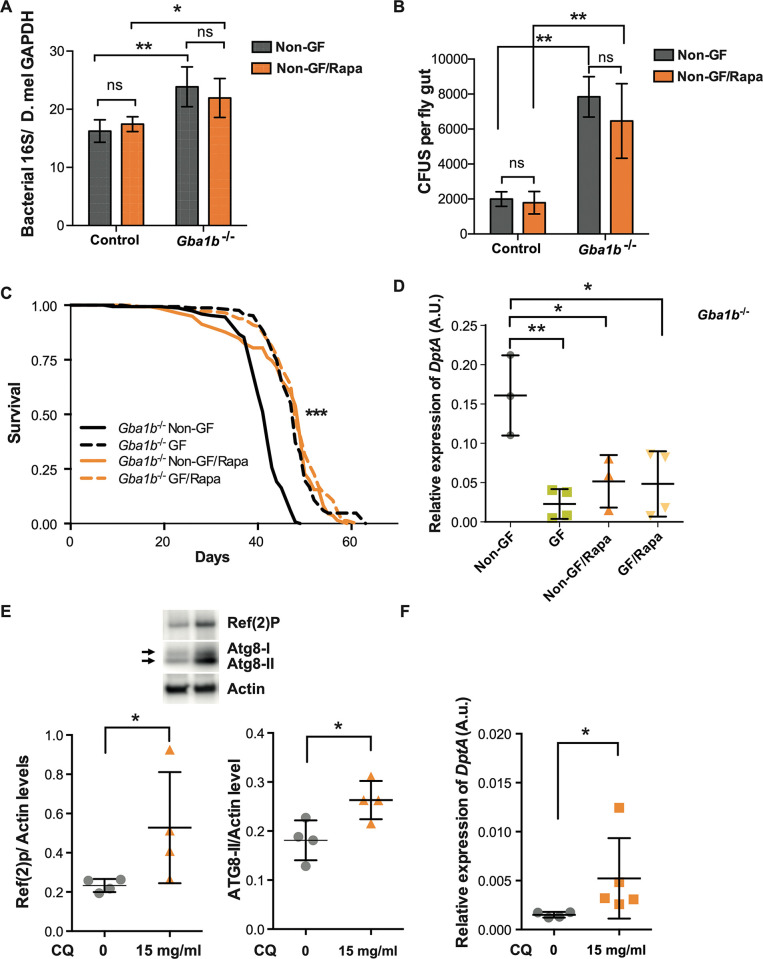
Mutations in the GBA1 gene cause the lysosomal storage disorder Gaucher disease (GD) and are the greatest known genetic risk factors for Parkinson's disease (PD). Communication between the gut and brain and immune dysregulation are increasingly being implicated in neurodegenerative disorders such as PD. Here, we show that flies lacking the Gba1b gene, the main fly orthologue of GBA1, display widespread NF-kB signalling activation, including gut inflammation, and brain glial activation. We also demonstrate intestinal autophagic defects, gut dysfunction, and microbiome dysbiosis. Remarkably, modulating the microbiome of Gba1b knockout flies, by raising them under germ-free conditions, partially ameliorates lifespan, locomotor and immune phenotypes. Moreover, we show that modulation of the immune deficiency (IMD) pathway is detrimental to the survival of Gba1 deficient flies. We also reveal that direct stimulation of autophagy by rapamycin treatment achieves similar benefits to germ-free conditions independent of gut bacterial load. Consistent with this, we show that pharmacologically blocking autophagosomal-lysosomal fusion, mimicking the autophagy defects of Gba1 depleted cells, is sufficient to stimulate intestinal immune activation. Overall, our data elucidate a mechanism whereby an altered microbiome, coupled with defects in autophagy, drive chronic activation of NF-kB signaling in a Gba1 loss-of-function model. It also highlights that elimination of the microbiota or stimulation of autophagy to remove immune mediators, rather than prolonged immunosuppression, may represent effective therapeutic avenues for GBA1-associated disorders.
Copyright: © 2023 Atilano et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Zunke F, Moise AC, Belur NR, Gelyana E, Stojkovska I, Dzaferbegovic H, et al.. Reversible Conformational Conversion of alpha-Synuclein into Toxic Assemblies by Glucosylceramide. Neuron. 2018;97(1):92–107 e10. Epub 2018/01/02. doi: 10.1016/j.neuron.2017.12.012 ; PubMed Central PMCID: PMC6013314. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
