

p53 deficient breast cancer cells reprogram preadipocytes toward tumor-protective immunomodulatory cells
- PMID: 38127986
- PMCID: PMC10756271
- DOI: 10.1073/pnas.2311460120
p53 deficient breast cancer cells reprogram preadipocytes toward tumor-protective immunomodulatory cells
Abstract
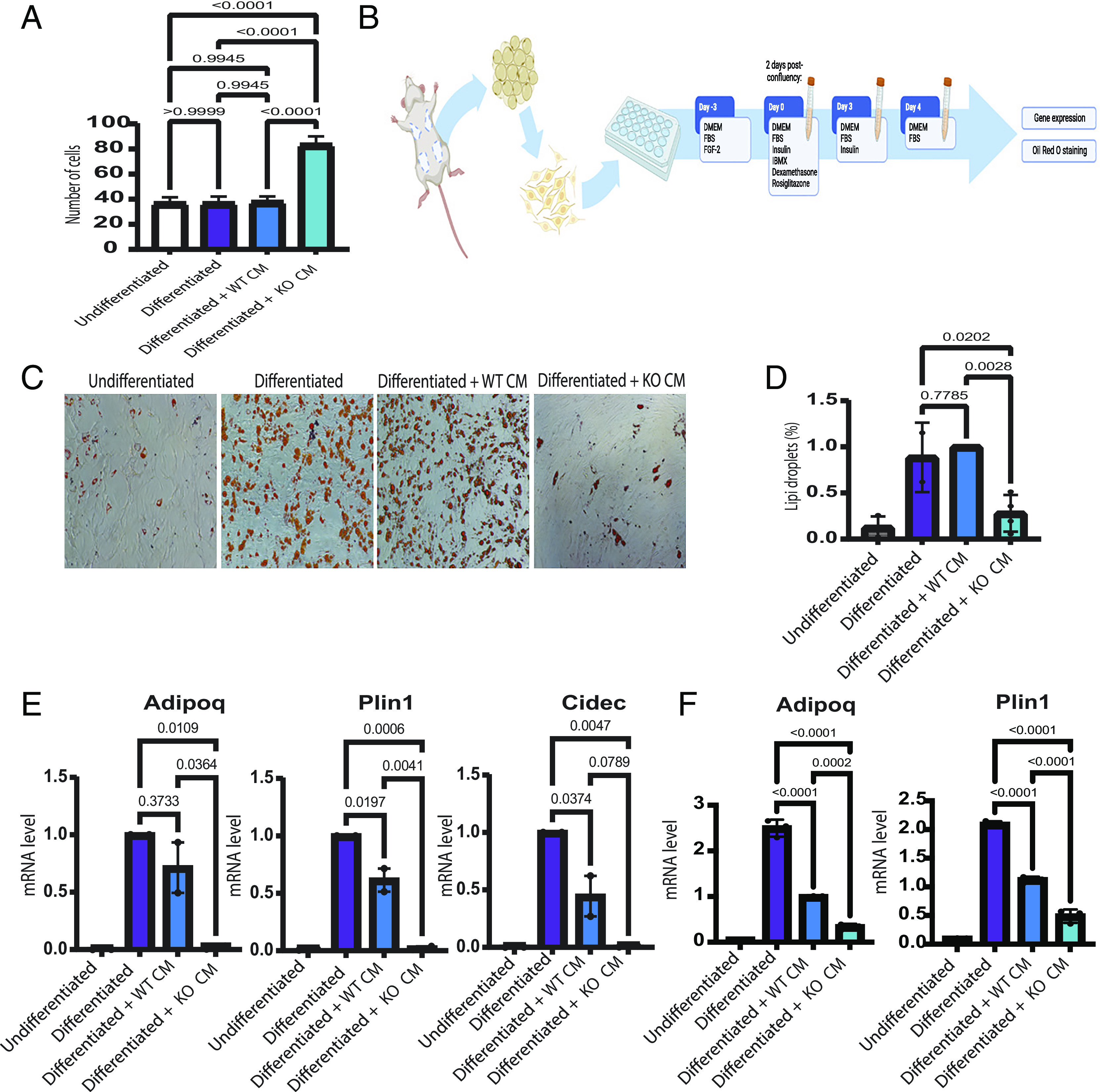
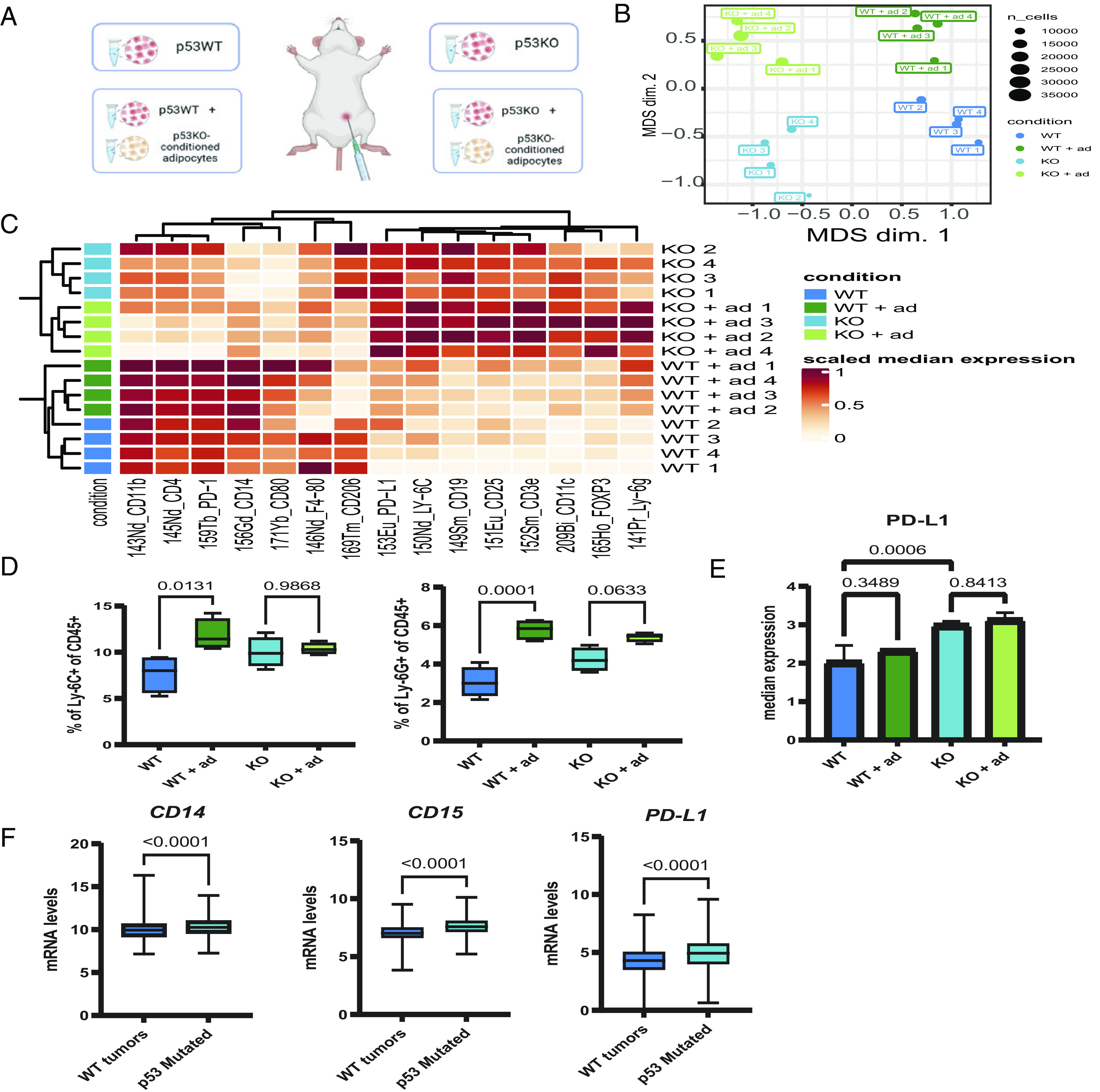
The TP53 gene is mutated in approximately 30% of all breast cancer cases. Adipocytes and preadipocytes, which constitute a substantial fraction of the stroma of normal mammary tissue and breast tumors, undergo transcriptional, metabolic, and phenotypic reprogramming during breast cancer development and play an important role in tumor progression. We report here that p53 loss in breast cancer cells facilitates the reprogramming of preadipocytes, inducing them to acquire a unique transcriptional and metabolic program that combines impaired adipocytic differentiation with augmented cytokine expression. This, in turn, promotes the establishment of an inflammatory tumor microenvironment, including increased abundance of Ly6C+ and Ly6G+ myeloid cells and elevated expression of the immune checkpoint ligand PD-L1. We also describe a potential gain-of-function effect of common p53 missense mutations on the inflammatory reprogramming of preadipocytes. Altogether, our study implicates p53 deregulation in breast cancer cells as a driver of tumor-supportive adipose tissue reprogramming, expanding the network of non-cell autonomous mechanisms whereby p53 dysfunction may promote cancer. Further elucidation of the interplay between p53 and adipocytes within the tumor microenvironment may suggest effective therapeutic targets for the treatment of breast cancer patients.
Keywords: adipocytes; breast cancer; p53; preadipocytes.
Conflict of interest statement
Competing interests statement:K.E.d.V. is consultant for Macomics.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
