

On the Origin and Evolution of the Mosquito Male-determining Factor Nix
- PMID: 38128148
- PMCID: PMC10798136
- DOI: 10.1093/molbev/msad276
On the Origin and Evolution of the Mosquito Male-determining Factor Nix
Abstract
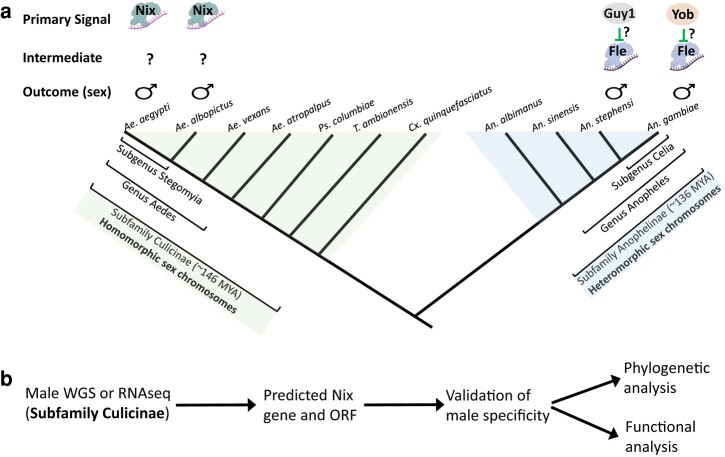
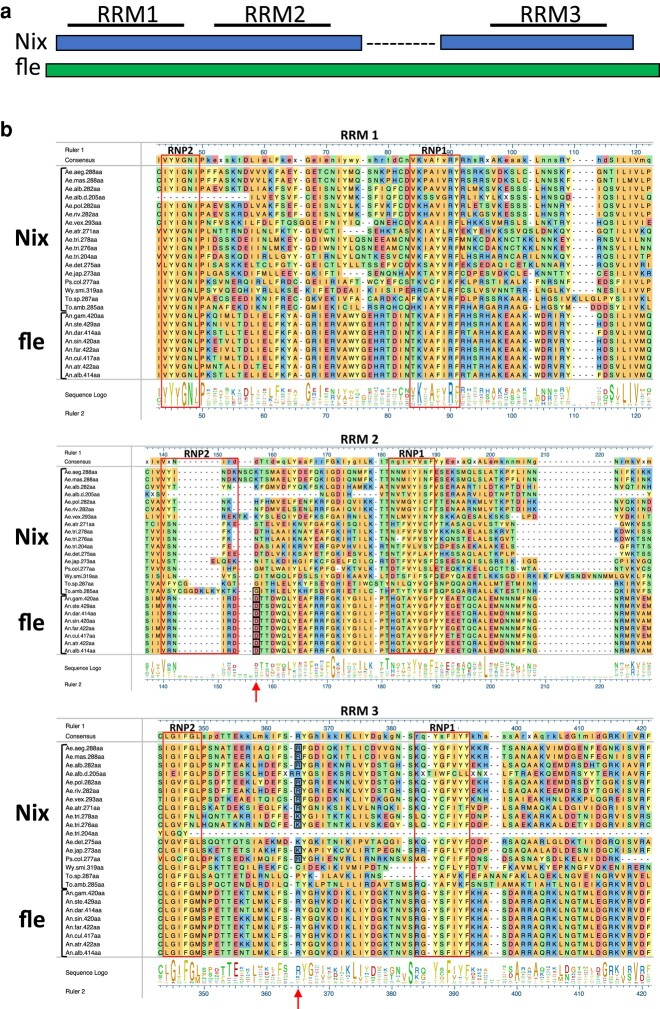
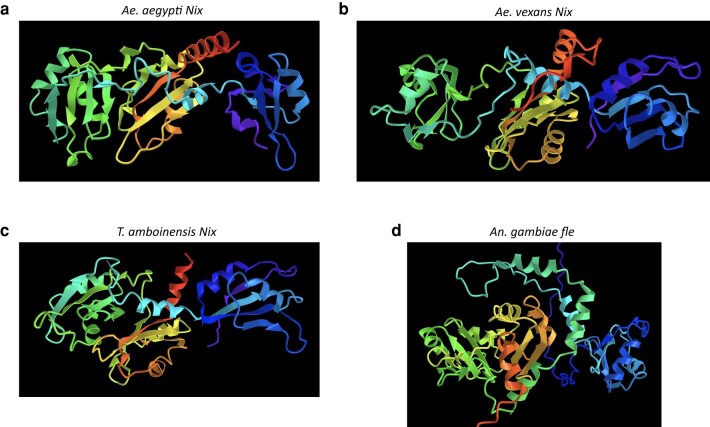
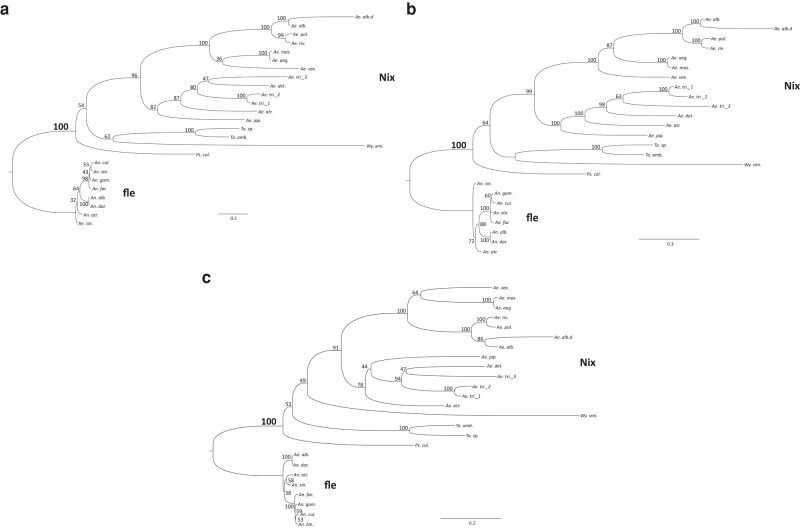
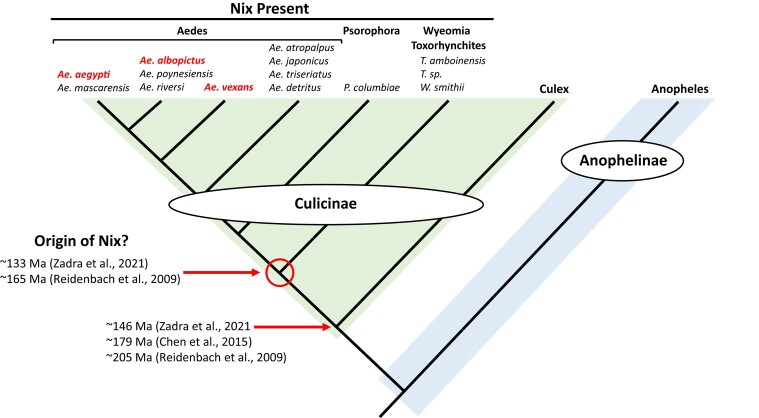
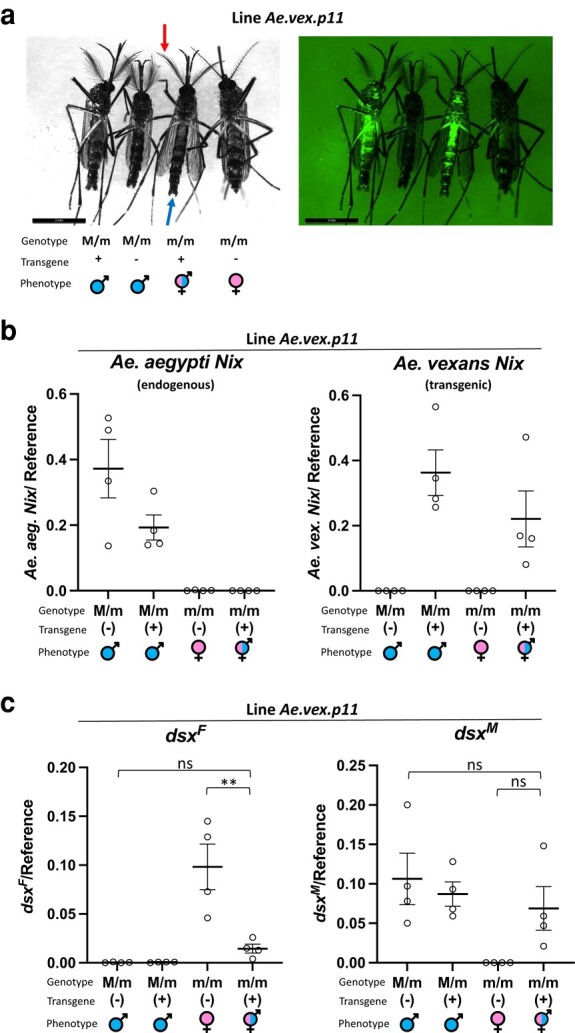
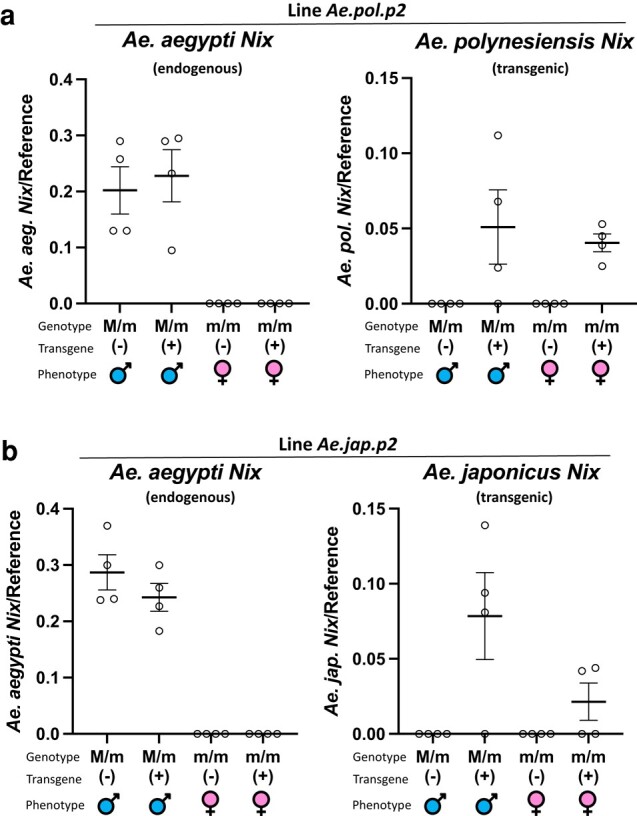
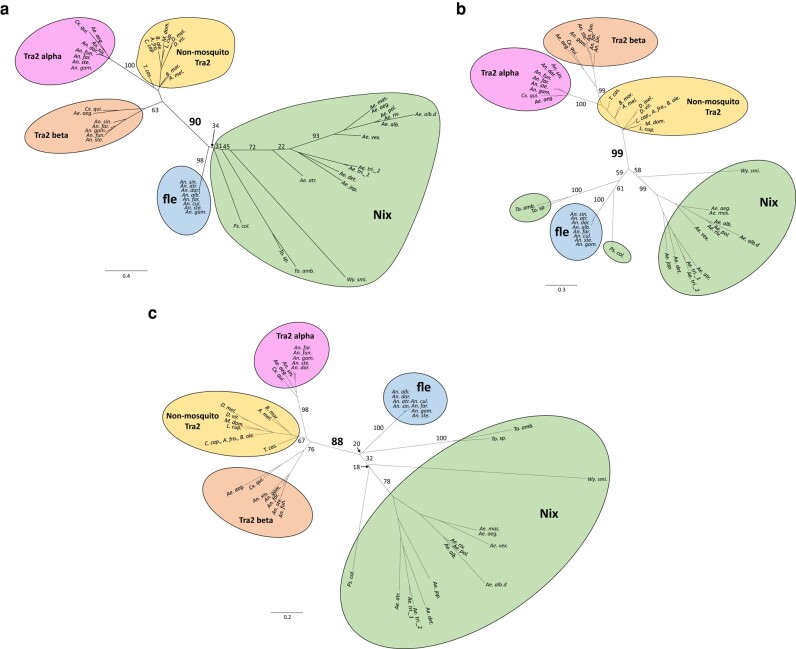
The mosquito family Culicidae is divided into 2 subfamilies named the Culicinae and Anophelinae. Nix, the dominant male-determining factor, has only been found in the culicines Aedes aegypti and Aedes albopictus, 2 important arboviral vectors that belong to the subgenus Stegomyia. Here we performed sex-specific whole-genome sequencing and RNAseq of divergent mosquito species and explored additional male-inclusive datasets to investigate the distribution of Nix. Except for the Culex genus, Nix homologs were found in all species surveyed from the Culicinae subfamily, including 12 additional species from 3 highly divergent tribes comprising 4 genera, suggesting Nix originated at least 133 to 165 million years ago (MYA). Heterologous expression of 1 of 3 divergent Nix open reading frames (ORFs) in Ae. aegypti resulted in partial masculinization of genetic females as evidenced by morphology and doublesex splicing. Phylogenetic analysis suggests Nix is related to femaleless (fle), a recently described intermediate sex-determining factor found exclusively in anopheline mosquitoes. Nix from all species has a conserved structure, including 3 RNA-recognition motifs (RRMs), as does fle. However, Nix has evolved at a much faster rate than fle. The RRM3 of both Nix and fle are distantly related to the single RRM of a widely distributed and conserved splicing factor transformer-2 (tra2). The RRM3-based phylogenetic analysis suggests this domain in Nix and fle may have evolved from tra2 or a tra2-related gene in a common ancestor of mosquitoes. Our results provide insights into the evolution of sex determination in mosquitoes and will inform broad applications of mosquito-control strategies based on manipulating sex ratios toward nonbiting males.
Keywords: genetic engineering; homomorphic sex chromosome; male-determining factor; mosquito control; sex determination.
© The Author(s) 2023. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures
Similar articles
-
Two of the three Transformer-2 genes are required for ovarian development in Aedes albopictus.Insect Biochem Mol Biol. 2019 Jun;109:92-105. doi: 10.1016/j.ibmb.2019.03.008. Epub 2019 Mar 23. Insect Biochem Mol Biol. 2019. PMID: 30914323 Free PMC article.
-
The Nix locus on the male-specific homologue of chromosome 1 in Aedes albopictus is a strong candidate for a male-determining factor.Parasit Vectors. 2018 Dec 24;11(Suppl 2):647. doi: 10.1186/s13071-018-3215-8. Parasit Vectors. 2018. PMID: 30583734 Free PMC article.
-
Nix is a male-determining factor in the Asian tiger mosquito Aedes albopictus.Insect Biochem Mol Biol. 2020 Mar;118:103311. doi: 10.1016/j.ibmb.2019.103311. Epub 2019 Dec 31. Insect Biochem Mol Biol. 2020. PMID: 31901476 Free PMC article.
-
Mosquitoes (Diptera: Culicidae) and mosquito-borne diseases in Mali, West Africa.Parasit Vectors. 2018 Aug 13;11(1):467. doi: 10.1186/s13071-018-3045-8. Parasit Vectors. 2018. PMID: 30103823 Free PMC article. Review.
-
Selective targeting of biting females to control mosquito-borne infectious diseases.Trends Parasitol. 2022 Sep;38(9):791-804. doi: 10.1016/j.pt.2022.05.012. Epub 2022 Jun 13. Trends Parasitol. 2022. PMID: 35952630 Free PMC article. Review.
Cited by
-
The chromosome-scale genome assembly for the West Nile vector Culex quinquefasciatus uncovers patterns of genome evolution in mosquitoes.BMC Biol. 2024 Jan 25;22(1):16. doi: 10.1186/s12915-024-01825-0. BMC Biol. 2024. PMID: 38273363 Free PMC article.
-
Hybridization between Aedes aegypti and Aedes mascarensis mosquitoes leads to disruption of male sex determination.Commun Biol. 2024 Jul 22;7(1):886. doi: 10.1038/s42003-024-06560-4. Commun Biol. 2024. PMID: 39039226 Free PMC article.
-
Mosquito sex determination: recent advances and applications.Curr Opin Insect Sci. 2025 Oct;71:101385. doi: 10.1016/j.cois.2025.101385. Epub 2025 May 13. Curr Opin Insect Sci. 2025. PMID: 40368280 Review.
References
-
- Aryan A, Anderson MAE, Biedler JK, Qi Y, Overcash JM, Naumenko AN, Sharakhova MV, Mao C, Adelman ZN, Tu Z. Nix alone is sufficient to convert female Aedes aegypti into fertile males and myo-sex is needed for male flight. Proc Natl Acad Sci U S A. 2020:117(30):17702–17709. 10.1073/pnas.2001132117. - DOI - PMC - PubMed
MeSH terms
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
