

A novel SMARCC1 BAFopathy implicates neural progenitor epigenetic dysregulation in human hydrocephalus
- PMID: 38128548
- PMCID: PMC10994532
- DOI: 10.1093/brain/awad405
A novel SMARCC1 BAFopathy implicates neural progenitor epigenetic dysregulation in human hydrocephalus
Abstract
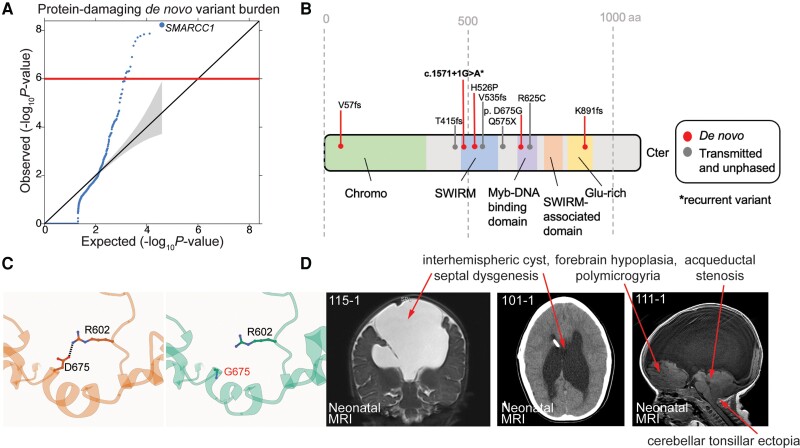
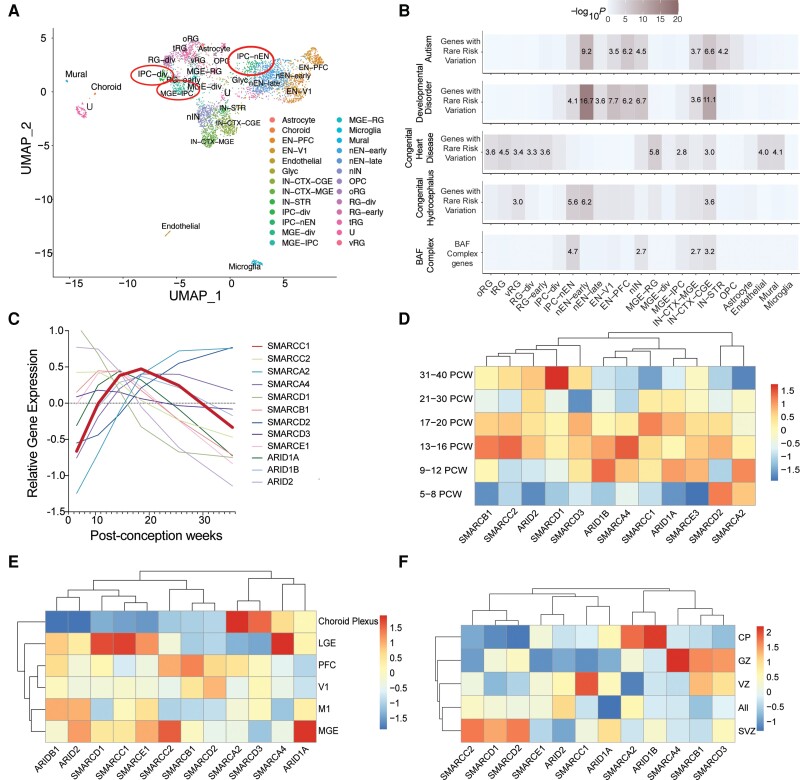
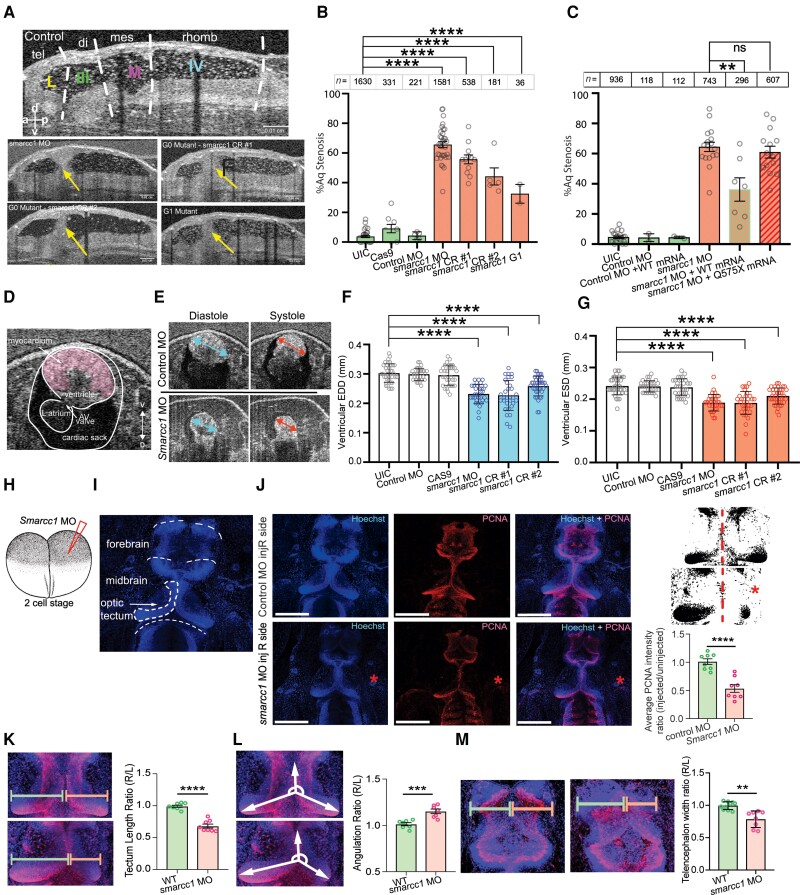
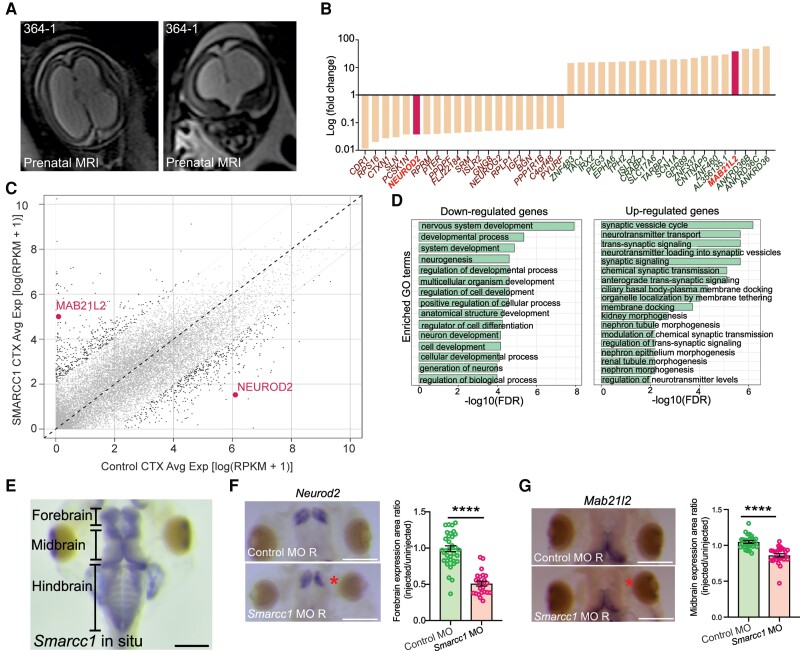
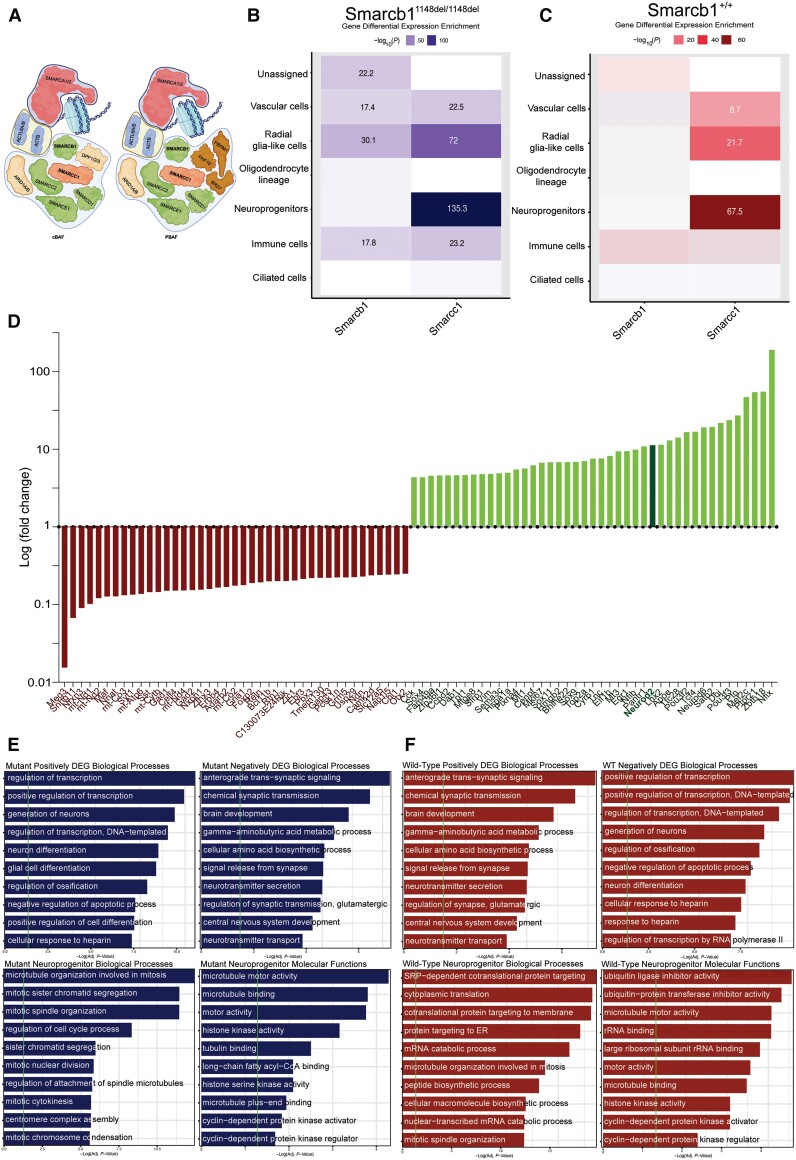
Hydrocephalus, characterized by cerebral ventriculomegaly, is the most common disorder requiring brain surgery in children. Recent studies have implicated SMARCC1, a component of the BRG1-associated factor (BAF) chromatin remodelling complex, as a candidate congenital hydrocephalus gene. However, SMARCC1 variants have not been systematically examined in a large patient cohort or conclusively linked with a human syndrome. Moreover, congenital hydrocephalus-associated SMARCC1 variants have not been functionally validated or mechanistically studied in vivo. Here, we aimed to assess the prevalence of SMARCC1 variants in an expanded patient cohort, describe associated clinical and radiographic phenotypes, and assess the impact of Smarcc1 depletion in a novel Xenopus tropicalis model of congenital hydrocephalus. To do this, we performed a genetic association study using whole-exome sequencing from a cohort consisting of 2697 total ventriculomegalic trios, including patients with neurosurgically-treated congenital hydrocephalus, that total 8091 exomes collected over 7 years (2016-23). A comparison control cohort consisted of 1798 exomes from unaffected siblings of patients with autism spectrum disorder and their unaffected parents were sourced from the Simons Simplex Collection. Enrichment and impact on protein structure were assessed in identified variants. Effects on the human fetal brain transcriptome were examined with RNA-sequencing and Smarcc1 knockdowns were generated in Xenopus and studied using optical coherence tomography imaging, in situ hybridization and immunofluorescence. SMARCC1 surpassed genome-wide significance thresholds, yielding six rare, protein-altering de novo variants localized to highly conserved residues in key functional domains. Patients exhibited hydrocephalus with aqueductal stenosis; corpus callosum abnormalities, developmental delay, and cardiac defects were also common. Xenopus knockdowns recapitulated both aqueductal stenosis and cardiac defects and were rescued by wild-type but not patient-specific variant SMARCC1. Hydrocephalic SMARCC1-variant human fetal brain and Smarcc1-variant Xenopus brain exhibited a similarly altered expression of key genes linked to midgestational neurogenesis, including the transcription factors NEUROD2 and MAB21L2. These results suggest de novo variants in SMARCC1 cause a novel human BAFopathy we term 'SMARCC1-associated developmental dysgenesis syndrome', characterized by variable presence of cerebral ventriculomegaly, aqueductal stenosis, developmental delay and a variety of structural brain or cardiac defects. These data underscore the importance of SMARCC1 and the BAF chromatin remodelling complex for human brain morphogenesis and provide evidence for a 'neural stem cell' paradigm of congenital hydrocephalus pathogenesis. These results highlight utility of trio-based whole-exome sequencing for identifying pathogenic variants in sporadic congenital structural brain disorders and suggest whole-exome sequencing may be a valuable adjunct in clinical management of congenital hydrocephalus patients.
Keywords: chromatin; congenital; genetics; genomics; neurodevelopment; neurosurgery.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For commercial re-use, please contact reprints@oup.com for reprints and translation rights for reprints. All other permissions can be obtained through our RightsLink service via the Permissions link on the article page on our site—for further information please contact journals.permissions@oup.com.
Conflict of interest statement
S.M. and P.K. are employees of GeneDx. All other co-authors have no conflicts of interest to disclose.
Figures
Update of
-
A novel SMARCC1 -mutant BAFopathy implicates epigenetic dysregulation of neural progenitors in hydrocephalus.medRxiv [Preprint]. 2023 Mar 20:2023.03.19.23287455. doi: 10.1101/2023.03.19.23287455. medRxiv. 2023. Update in: Brain. 2024 Apr 4;147(4):1553-1570. doi: 10.1093/brain/awad405. PMID: 36993720 Free PMC article. Updated. Preprint.
References
-
- Masliah-Planchon J, Bieche I, Guinebretiere JM, Bourdeaut F, Delattre O. SWI/SNF chromatin remodeling and human malignancies. Annu Rev Pathol. 2015;10:145–171. - PubMed
-
- Clapier CR, Cairns BR. The biology of chromatin remodeling complexes. Annu Rev Biochem. 2009;78:273–304. - PubMed
-
- Wang W, Xue Y, Zhou S, Kuo A, Cairns BR, Crabtree GR. Diversity and specialization of mammalian SWI/SNF complexes. Genes Dev. 1996;10:2117–2130. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- R01 NS109358/NS/NINDS NIH HHS/United States
- R01 HD104938/HD/NICHD NIH HHS/United States
- R01 NS111029/NS/NINDS NIH HHS/United States
- R01 NS127879/NS/NINDS NIH HHS/United States
- R00 HL143036/HL/NHLBI NIH HHS/United States
- U54 HG006504/HG/NHGRI NIH HHS/United States
- R21 NS116484/NS/NINDS NIH HHS/United States
- R01 NS111029-01A1/NH/NIH HHS/United States
- UM1 HG006493/HG/NHGRI NIH HHS/United States
- U24 HG008956/HG/NHGRI NIH HHS/United States
- R01 HL109942/HL/NHLBI NIH HHS/United States
- R00HL143036-02/HL/NHLBI NIH HHS/United States
- R01 NS131610/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
