

Chromatin regulator SMARCAL1 modulates cellular lipid metabolism
- PMID: 38129665
- PMCID: PMC10739977
- DOI: 10.1038/s42003-023-05665-6
Chromatin regulator SMARCAL1 modulates cellular lipid metabolism
Abstract
Biallelic mutations of the chromatin regulator SMARCAL1 cause Schimke Immunoosseous Dysplasia (SIOD), characterized by severe growth defects and premature mortality. Atherosclerosis and hyperlipidemia are common among SIOD patients, yet their onset and progression are poorly understood. Using an integrative approach involving proteomics, mouse models, and population genetics, we investigated SMARCAL1's role. We found that SmarcAL1 interacts with angiopoietin-like 3 (Angptl3), a key regulator of lipoprotein metabolism. In vitro and in vivo analyses demonstrate SmarcAL1's vital role in maintaining cellular lipid homeostasis. The observed translocation of SmarcAL1 to cytoplasmic peroxisomes suggests a potential regulatory role in lipid metabolism through gene expression. SmarcAL1 gene inactivation reduces the expression of key genes in cellular lipid catabolism. Population genetics investigations highlight significant associations between SMARCAL1 genetic variations and body mass index, along with lipid-related traits. This study underscores SMARCAL1's pivotal role in cellular lipid metabolism, likely contributing to the observed lipid phenotypes in SIOD patients.
© 2023. The Author(s).
Conflict of interest statement
The authors declare the following competing interests: D.J.R. serves on scientific advisory boards for Alnylam, Novartis, Pfizer, and Verve. C.A.C. is a founder and Chief Scientific Officer of Sana Biotechnology. The other authors declare no competing interests.
Figures


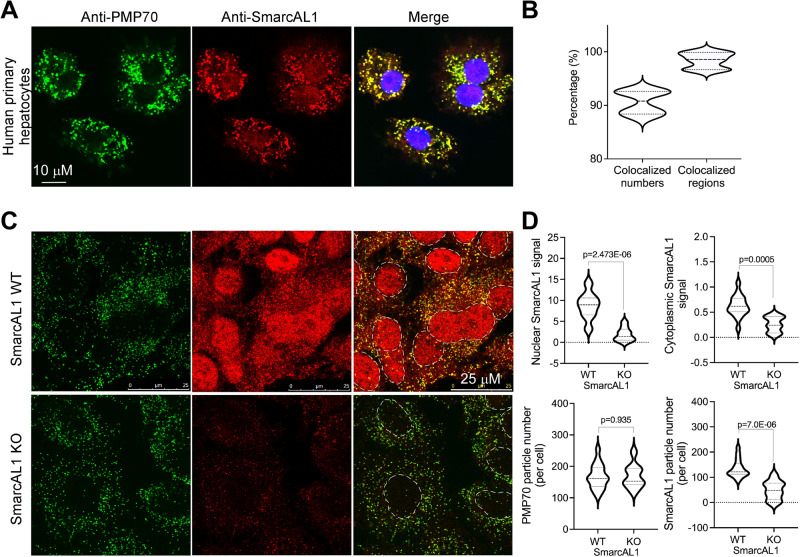

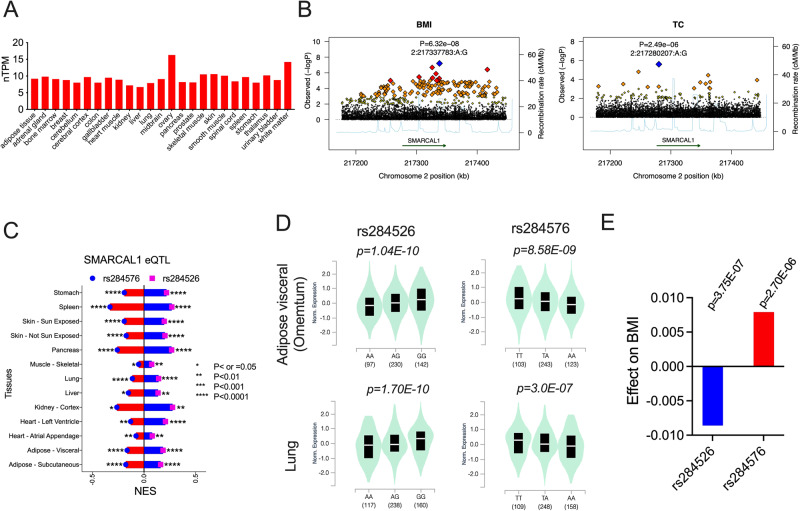
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
