

Schistosome egg-derived extracellular vesicles deliver Sja-miR-71a inhibits host macrophage and neutrophil extracellular traps via targeting Sema4D
- PMID: 38129877
- PMCID: PMC10734185
- DOI: 10.1186/s12964-023-01395-8
Schistosome egg-derived extracellular vesicles deliver Sja-miR-71a inhibits host macrophage and neutrophil extracellular traps via targeting Sema4D
Abstract
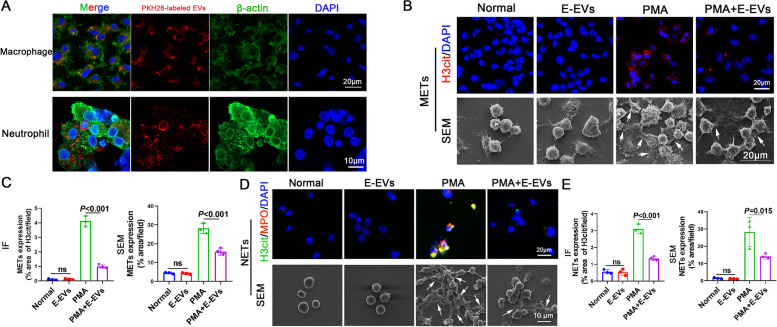
Background: Macrophages and neutrophils are rapidly recruited around Schistosome eggs to form granulomas. Extracellular traps (ETs) of macrophages and neutrophils are part of the pathogen clearance armamentarium of leukocytes. Schistosome eggs possess the ability to resist attack by the host's immune cells and survive by employing various immune evasion mechanisms, including the release of extracellular vesicles (EVs). However, the specific mechanisms by which Schistosome egg-derived EVs (E-EVs) evade the immune response and resist attack from macrophage and neutrophil ETs remain poorly understood. In this study, we aimed to investigate the association between E-EVs and macrophage/neutrophil ETs.
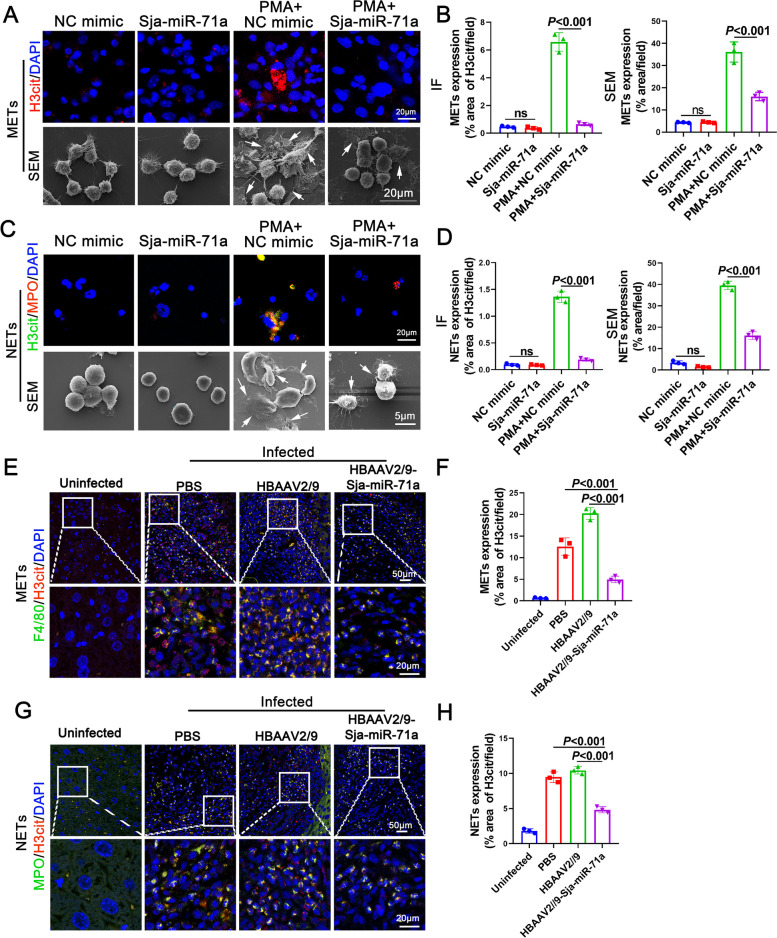
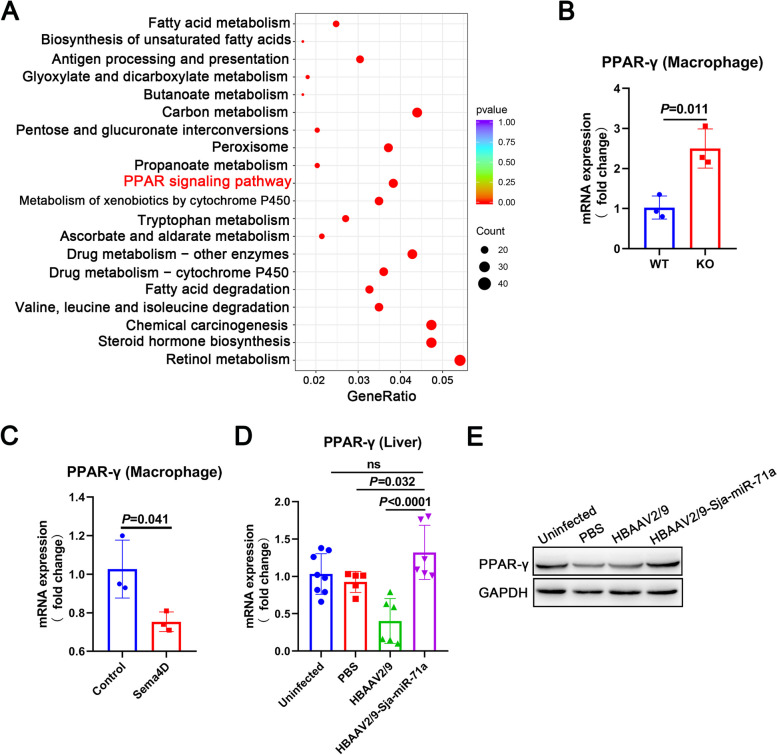
Methods: EVs were isolated from the culture supernatant of S. japonicum eggs and treated macrophages and neutrophils with E-EVs and Sja-miR-71a. The formation of ETs was then observed. Additionally, we infected mice with S. japonicum, administered HBAAV2/9-Sja-miR-71a, and the formation of macrophage ETs (METs) and neutrophil ETs (NETs) in the livers was measured. Sema4D-knockout mice, RNA sequencing, and trans-well assay were used to clarify Sja-miR-71a in E-EVs inhibits METs and NETs formation via the Sema4D/ PPAR-γ/ IL-10 axis.
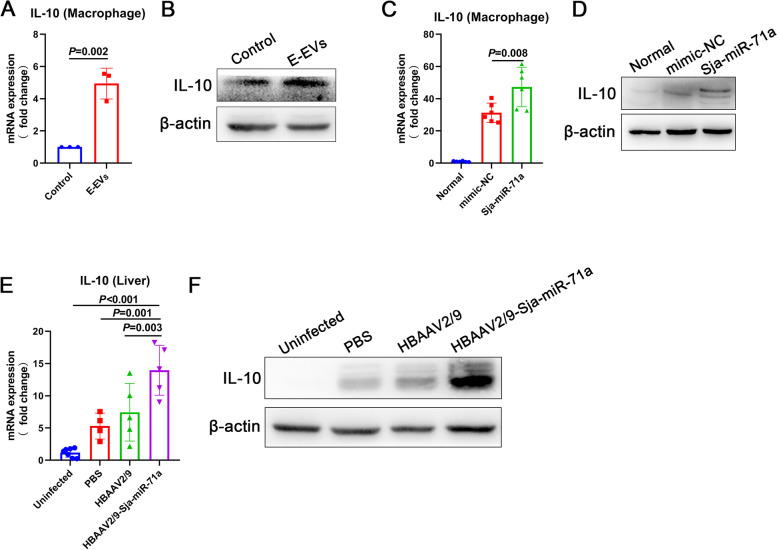
Results: Our findings revealed that E-EVs were internalized by macrophages and neutrophils, leading to the inhibition of METs and NETs formation. The highly expressed Sja-miR-71a in E-EVs targeted Sema4D, resulting in the up-regulation of IL-10 and subsequent inhibition of METs and NETs formation. Sema4D knockout up-regulated IL-10 expression and inhibited the formation of METs and NETs. Furthermore, we further demonstrated that Sja-miR-71a inhibits METs and NETs formation via the Sema4D/ PPAR-γ/ IL-10 axis.
Conclusions: In summary, our findings provide new insights into the immune evasion abilities of Schistosome eggs by demonstrating their ability to inhibit the formation of METs and NETs through the secretion of EVs. This study enhances our understanding of the host-pathogen interaction and may have implications for the development of novel therapeutic approaches. Video Abstract.
Keywords: Extracellular traps; Extracellular vesicles; Schistosoma japonicum eggs; Sema4D; Sja-miR-71a.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
