

Age-Related Alternative Splicing: Driver or Passenger in the Aging Process?
- PMID: 38132139
- PMCID: PMC10742131
- DOI: 10.3390/cells12242819
Age-Related Alternative Splicing: Driver or Passenger in the Aging Process?
Abstract
Alternative splicing changes are closely linked to aging, though it remains unclear if they are drivers or effects. As organisms age, splicing patterns change, varying gene isoform levels and functions. These changes may contribute to aging alterations rather than just reflect declining RNA quality control. Three main splicing types-intron retention, cassette exons, and cryptic exons-play key roles in age-related complexity. These events modify protein domains and increase nonsense-mediated decay, shifting protein isoform levels and functions. This may potentially drive aging or serve as a biomarker. Fluctuations in splicing factor expression also occur with aging. Somatic mutations in splicing genes can also promote aging and age-related disease. The interplay between splicing and aging has major implications for aging biology, though differentiating correlation and causation remains challenging. Declaring a splicing factor or event as a driver requires comprehensive evaluation of the associated molecular and physiological changes. A greater understanding of how RNA splicing machinery and downstream targets are impacted by aging is essential to conclusively establish the role of splicing in driving aging, representing a promising area with key implications for understanding aging, developing novel therapeutical options, and ultimately leading to an increase in the healthy human lifespan.
Keywords: age-associated splicing events; age-related diseases; aging; alternative splicing; molecular aging; senescence; splice variants; splicing factors; splicing regulation; transcriptome changes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
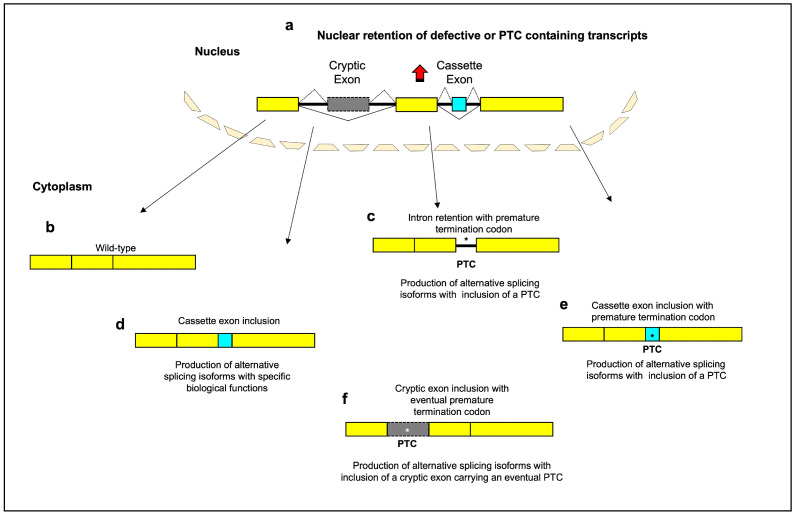
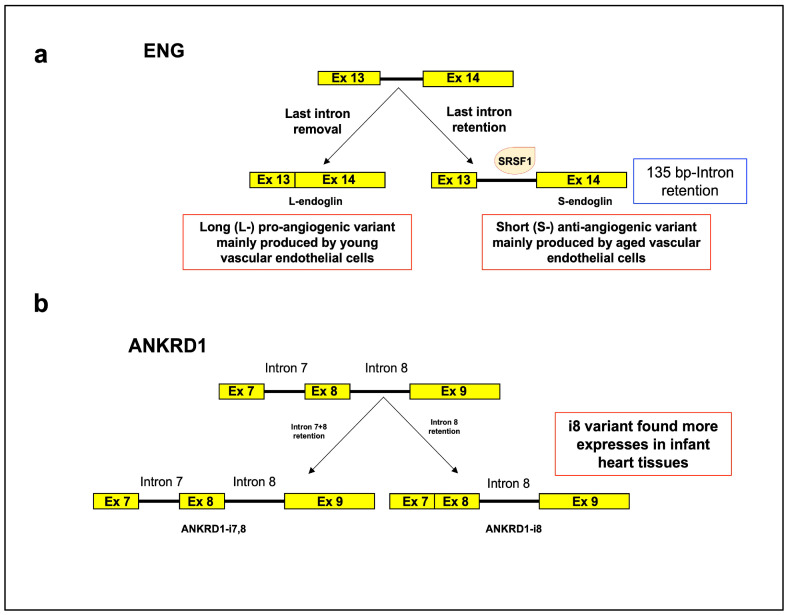
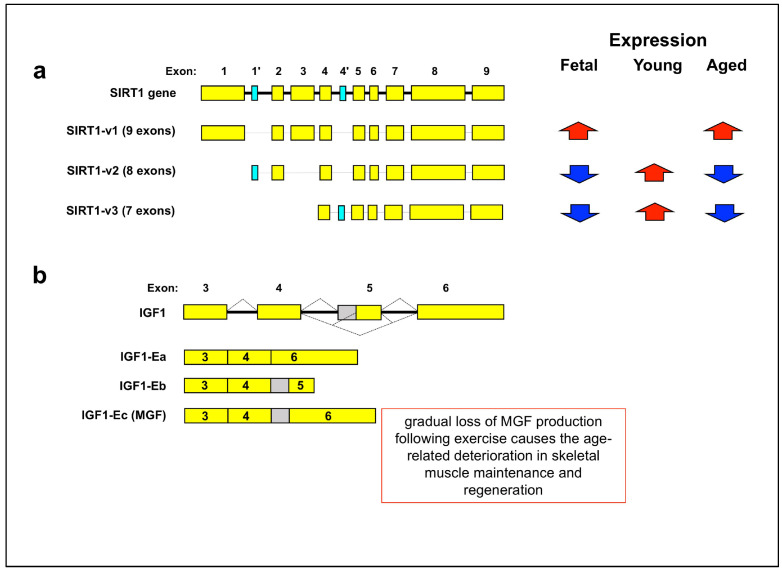
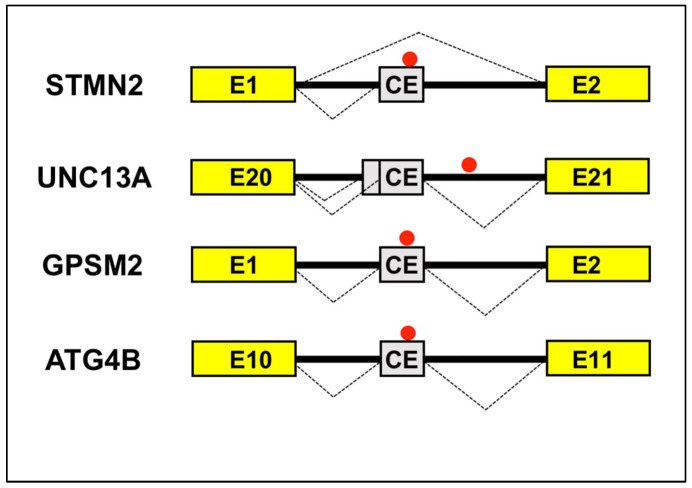
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
