

Spatially coordinated heterochromatinization of long synaptic genes in fragile X syndrome
- PMID: 38134876
- PMCID: PMC10794044
- DOI: 10.1016/j.cell.2023.11.019
Spatially coordinated heterochromatinization of long synaptic genes in fragile X syndrome
Abstract
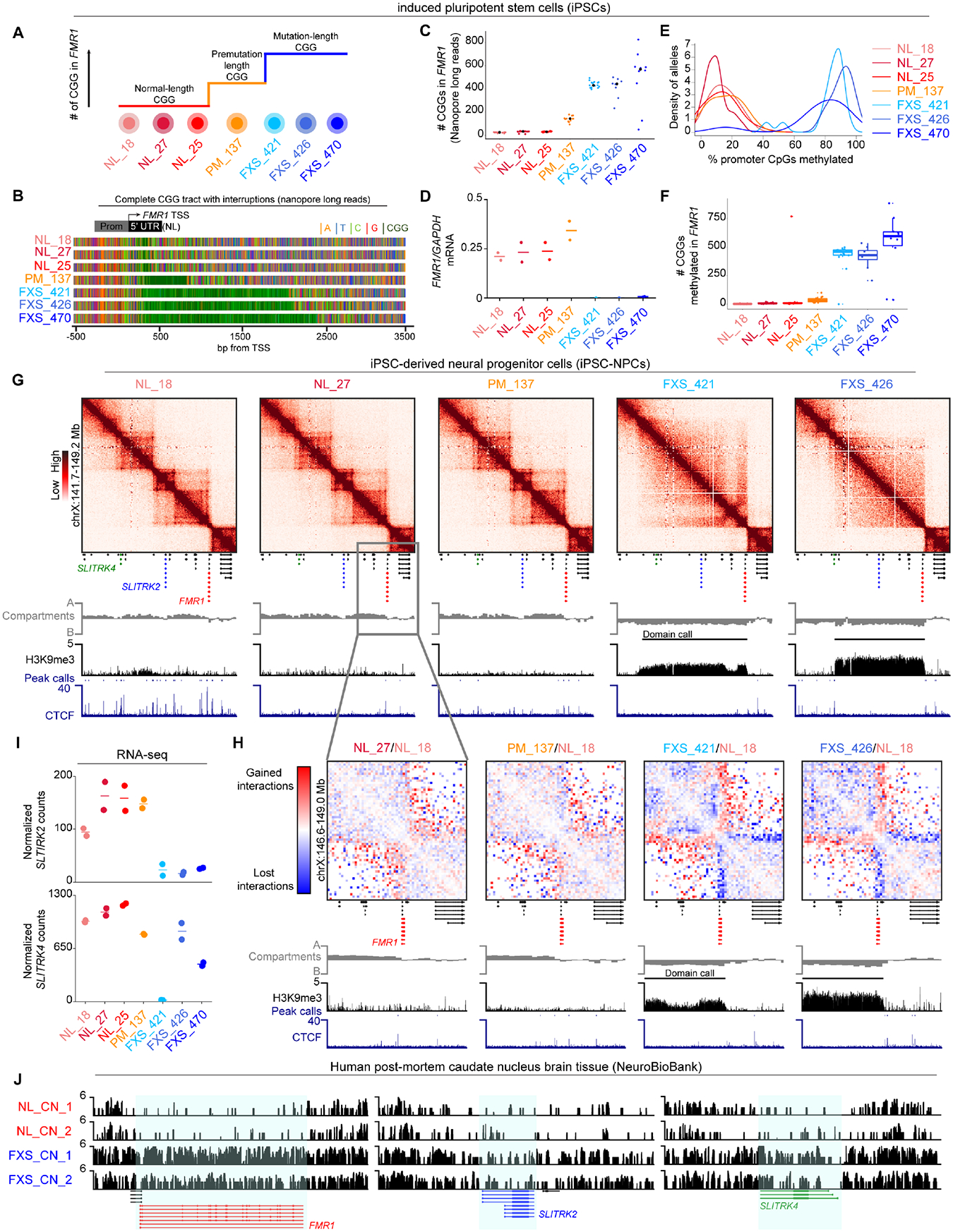
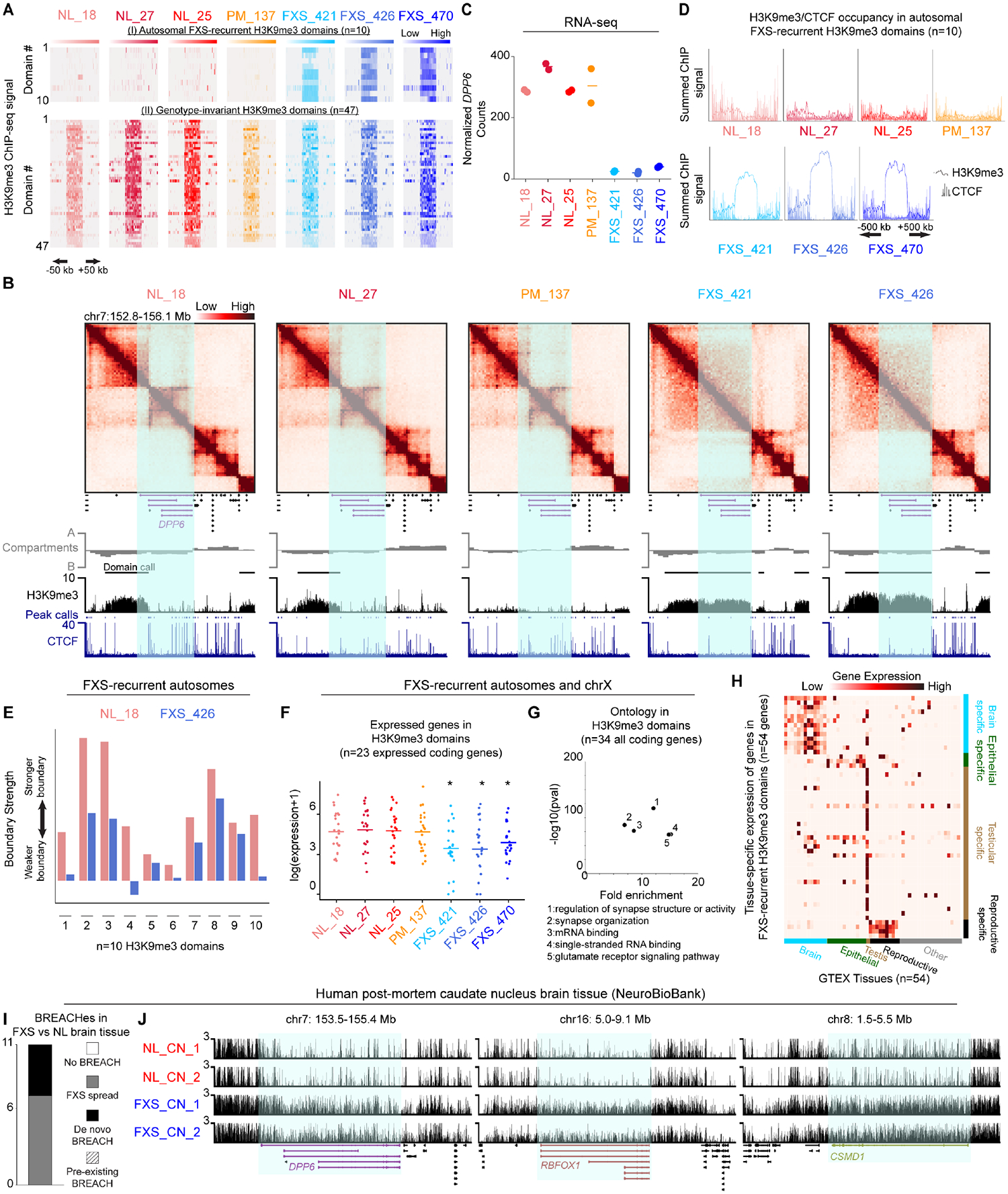
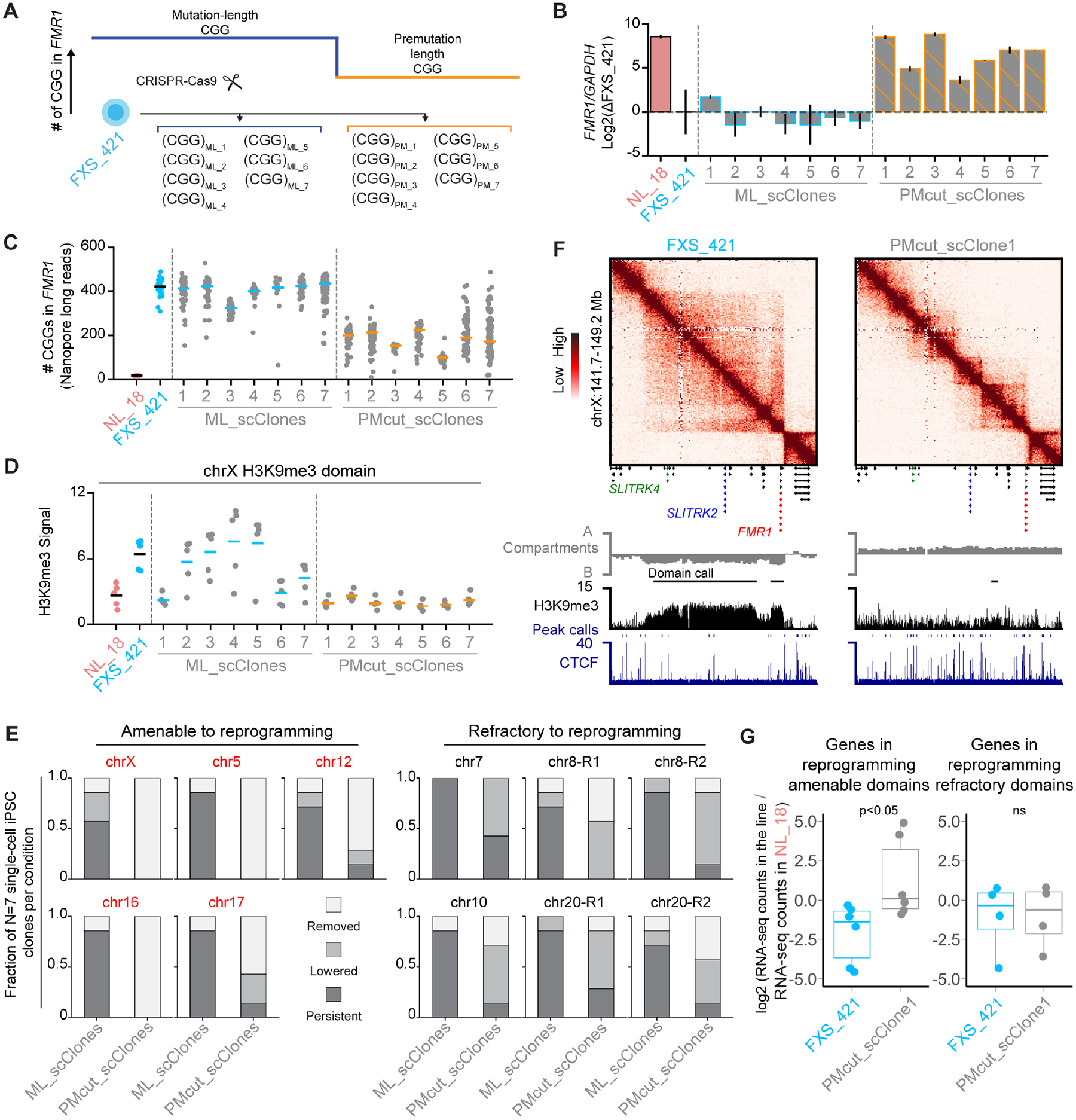
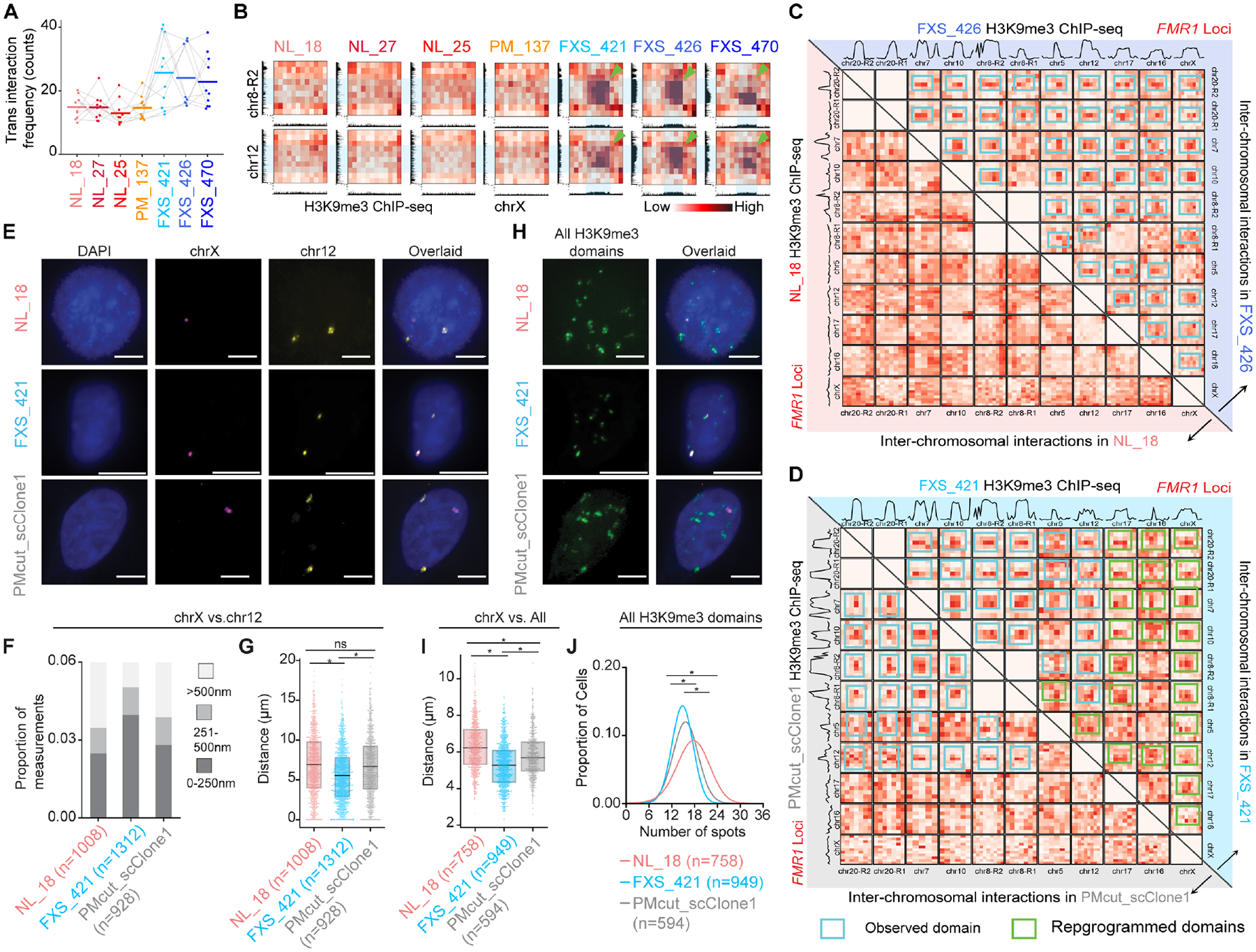
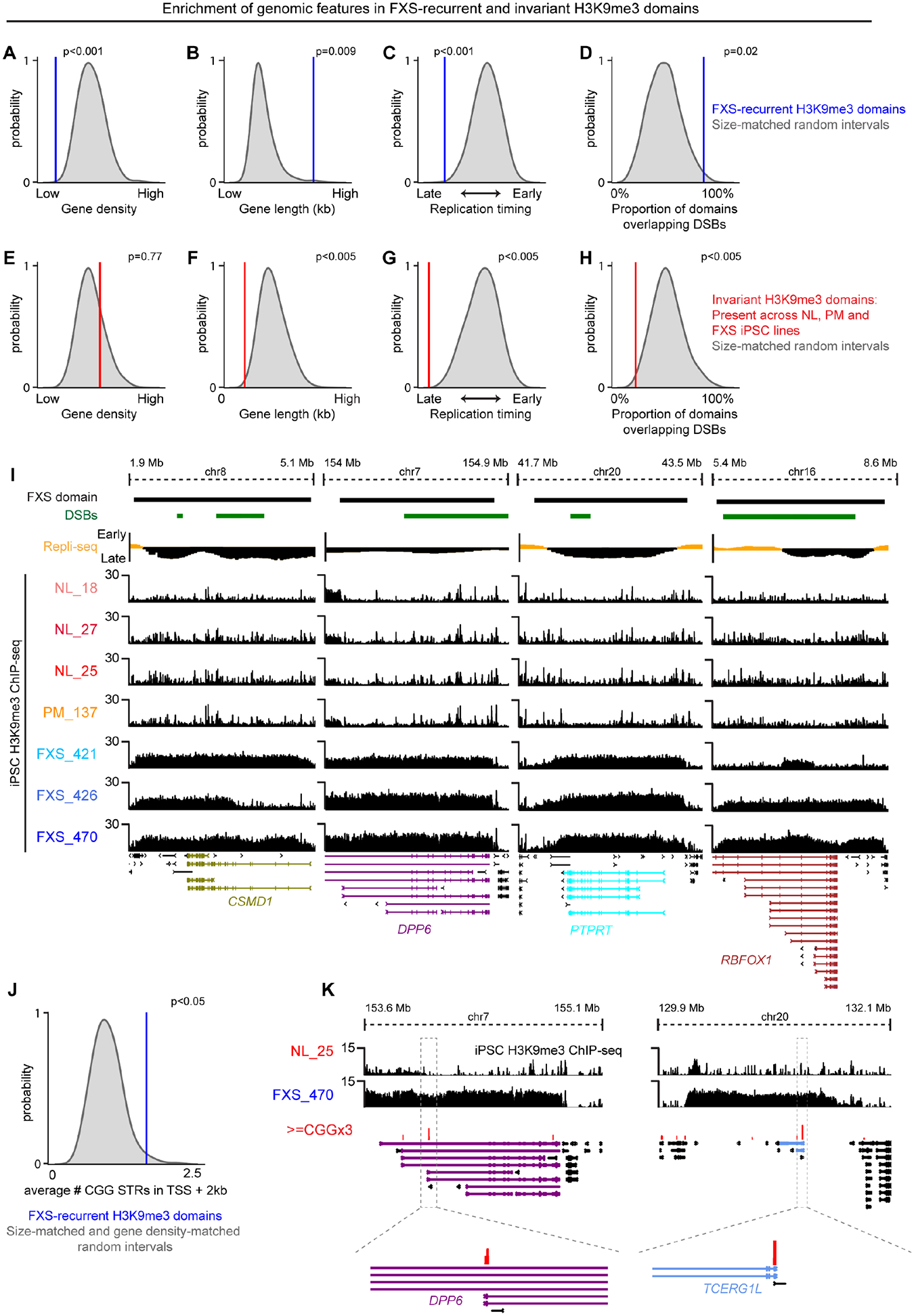
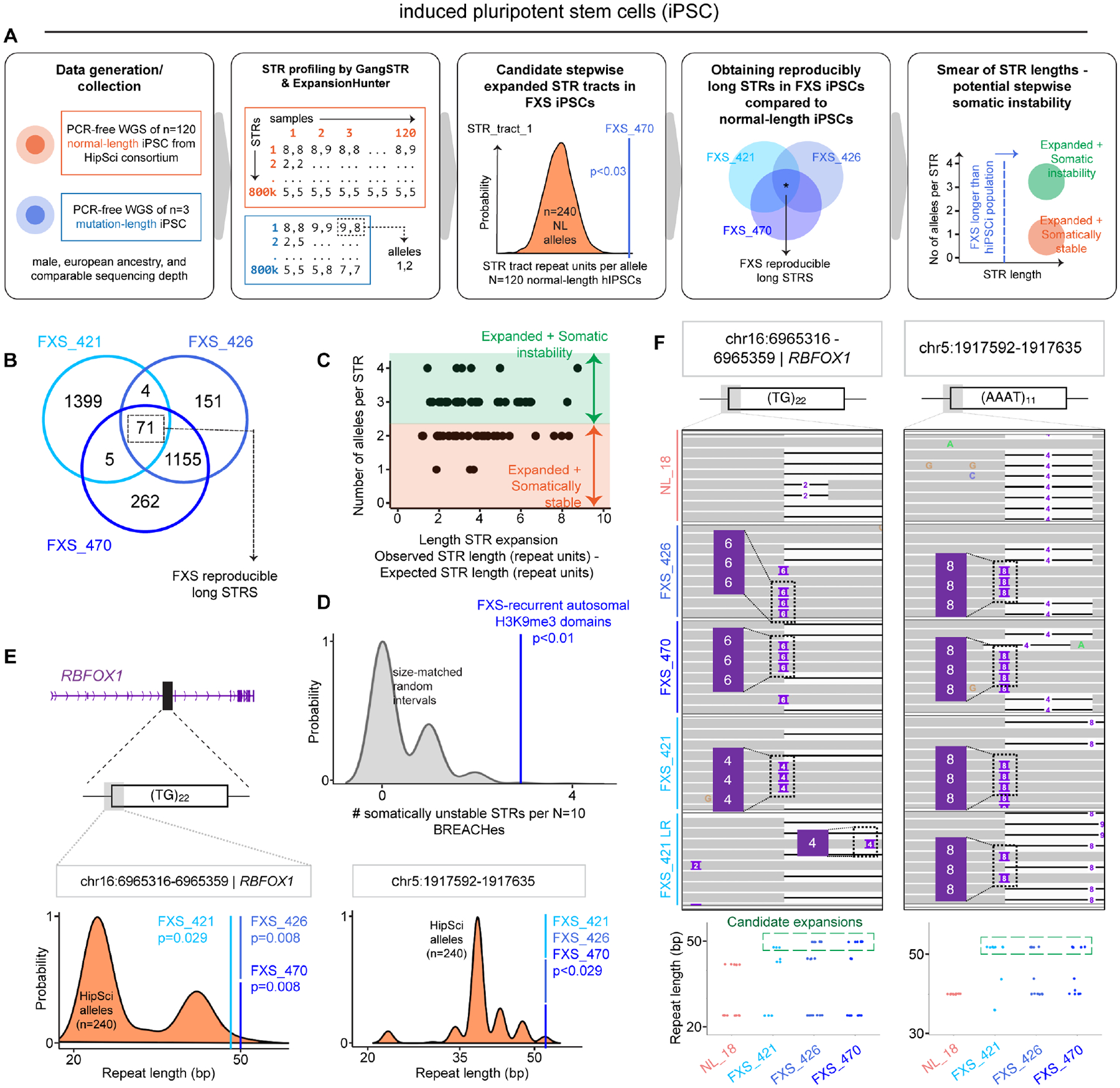
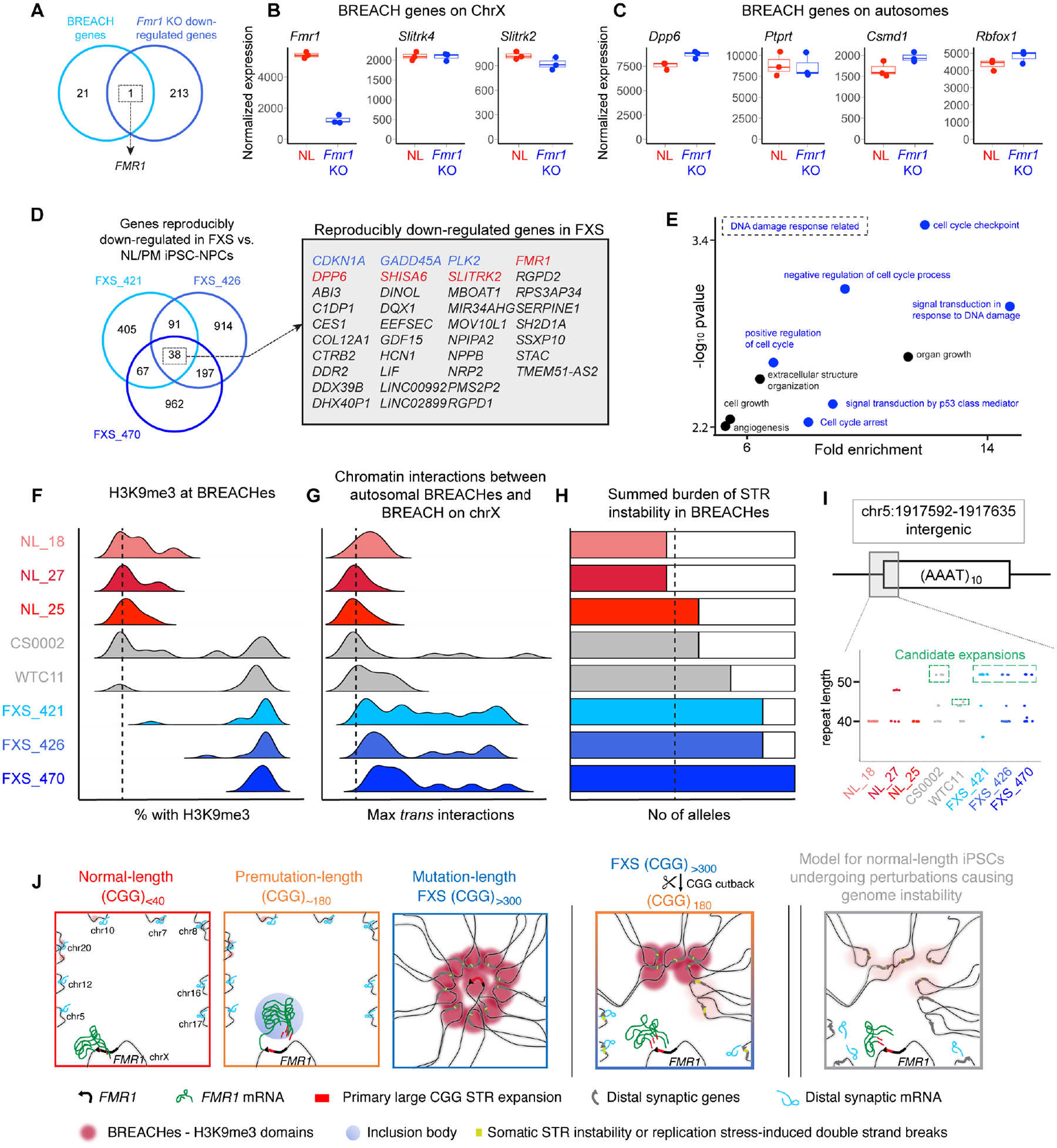
Short tandem repeat (STR) instability causes transcriptional silencing in several repeat expansion disorders. In fragile X syndrome (FXS), mutation-length expansion of a CGG STR represses FMR1 via local DNA methylation. Here, we find megabase-scale H3K9me3 domains on autosomes and encompassing FMR1 on the X chromosome in FXS patient-derived iPSCs, iPSC-derived neural progenitors, EBV-transformed lymphoblasts, and brain tissue with mutation-length CGG expansion. H3K9me3 domains connect via inter-chromosomal interactions and demarcate severe misfolding of TADs and loops. They harbor long synaptic genes replicating at the end of S phase, replication-stress-induced double-strand breaks, and STRs prone to stepwise somatic instability. CRISPR engineering of the mutation-length CGG to premutation length reverses H3K9me3 on the X chromosome and multiple autosomes, refolds TADs, and restores gene expression. H3K9me3 domains can also arise in normal-length iPSCs created with perturbations linked to genome instability, suggesting their relevance beyond FXS. Our results reveal Mb-scale heterochromatinization and trans interactions among loci susceptible to instability.
Keywords: CRISPR; DNA FISH; Hi-C; chromatin; epigenetics; fragile X syndrome; heterochromatin; higher-order genome folding; repeat expansion disorders; short tandem repeats; topologically associating domains.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests L.Z., C.G., and J.E.P.-C. are inventors on patent US20220323553A1 related to this work (https://patents.google.com/patent/US20220323553A1/en).
Figures
References
-
- Verkerk AJ, Pieretti M, Sutcliffe JS, Fu YH, Kuhl DP, Pizzuti A, Reiner O, Richards S, Victoria MF, Zhang FP, and et al. (1991). Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell 65, 905–914. 10.1016/0092-8674(91)90397-h. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
