

IDF-11774 Induces Cell Cycle Arrest and Apoptosis by Inhibiting HIF-1α in Gastric Cancer
- PMID: 38140111
- PMCID: PMC10748108
- DOI: 10.3390/pharmaceutics15122772
IDF-11774 Induces Cell Cycle Arrest and Apoptosis by Inhibiting HIF-1α in Gastric Cancer
Abstract
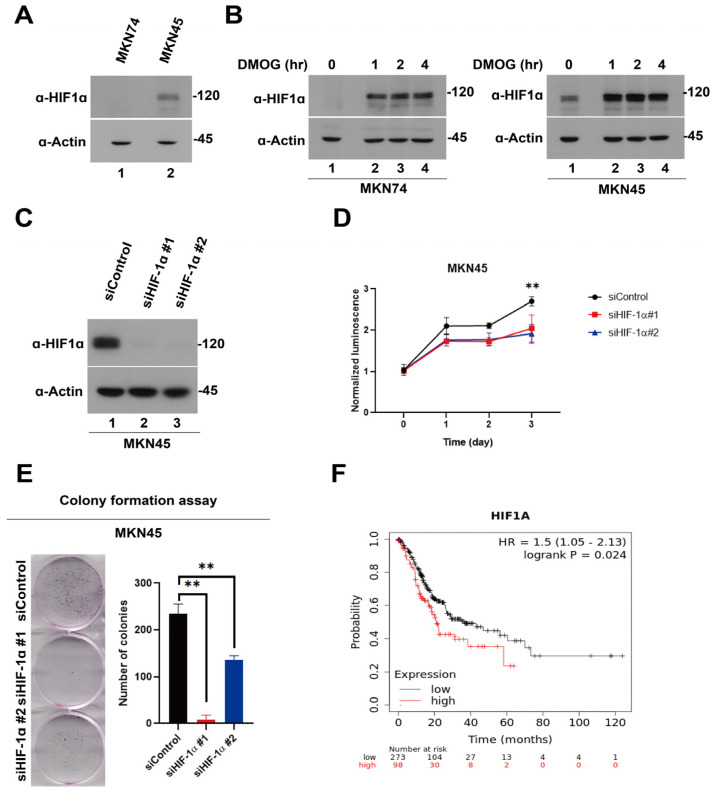
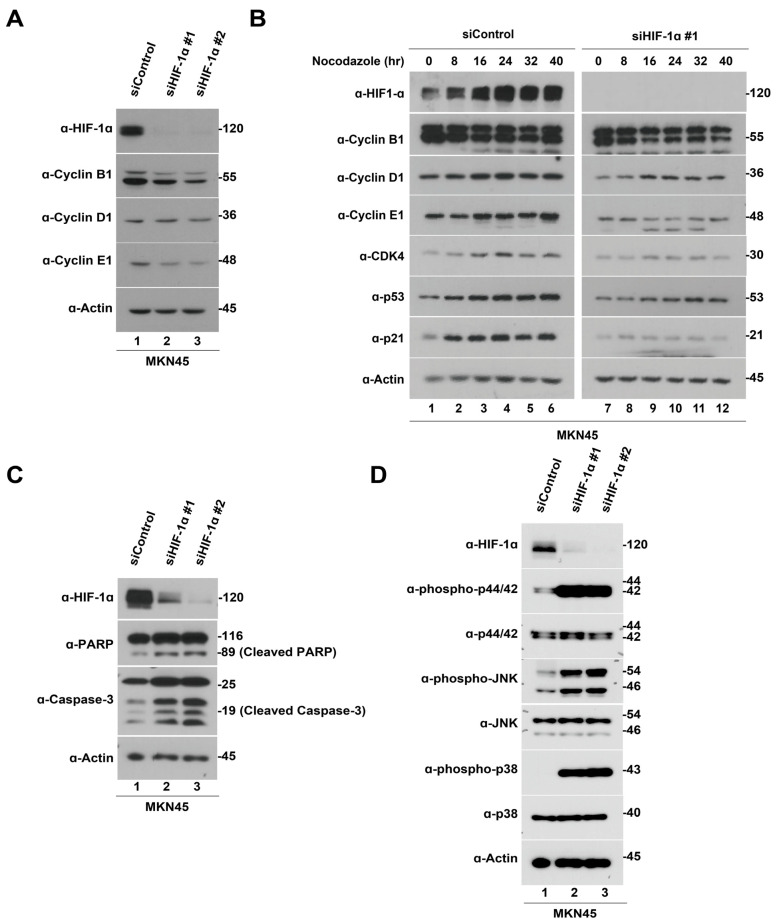
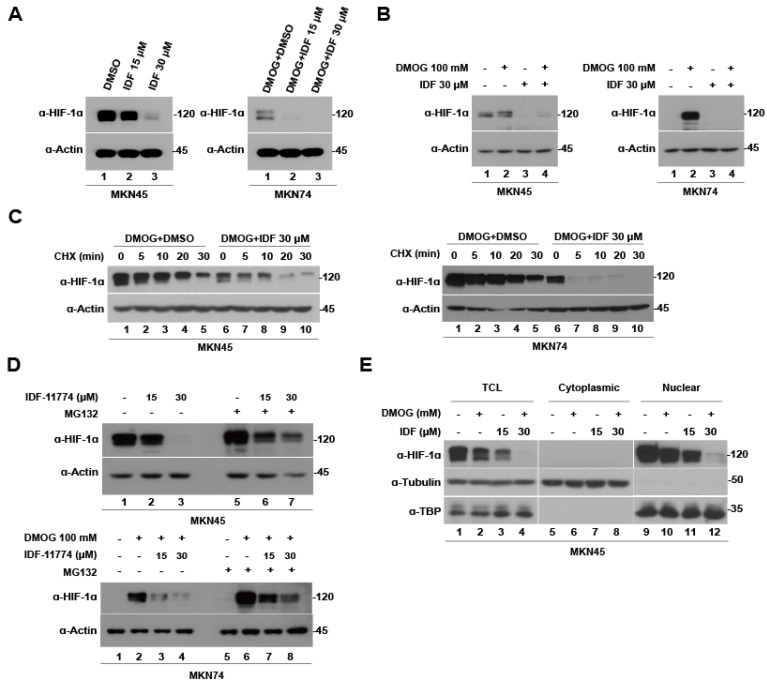
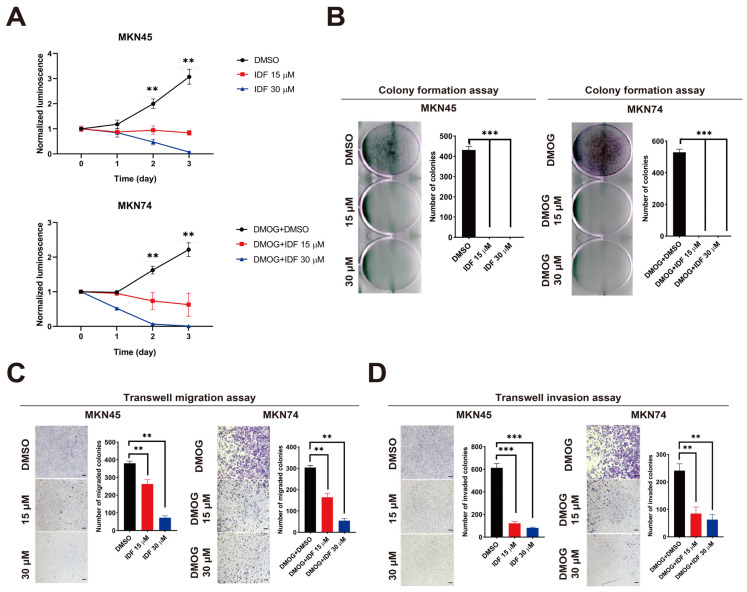
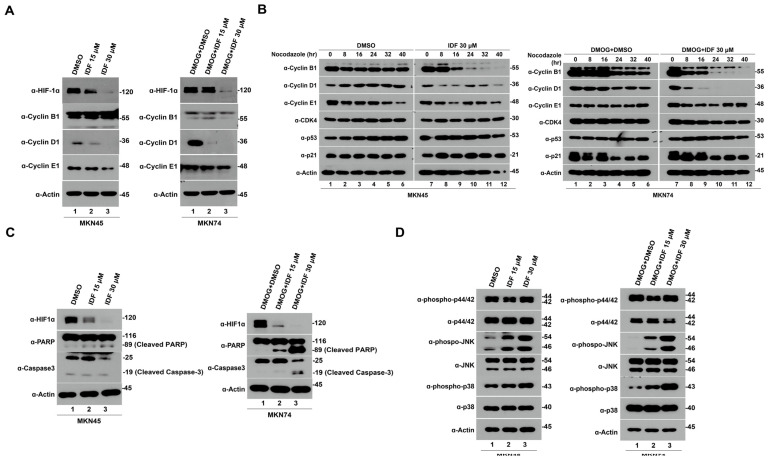
Hypoxia-inducible factor-1 alpha (HIF-1α) is a regulatory factor of intracellular oxygen supersession. The expression or increased activity of HIF-1α is closely related to various human cancers. Previously, IDF-11774 was demonstrated to inhibit HSP70 chaperone activity and suppress the accumulation of HIF-1α. In this study, we aimed to determine the effects of IDF-11774 on gastric cancer cell lines. Treatment with IDF-11774 was found to markedly decrease the proliferation, migration, and invasion of the gastric cancer cell lines. Furthermore, the phosphorylation levels of extracellular signal-regulated kinase 1/2, p38, and Jun N-terminal kinase in the mitogen-activated protein kinase signaling pathways were markedly increased in a dose-dependent manner, ultimately promoting apoptosis via the induction of cell cycle arrest. Our findings indicate that HIF-1α inhibitors are potent drugs for the treatment of gastric cancer.
Keywords: HIF-1α; HIF-1α inhibitor; IDF-11774; apoptosis; cell cycle arrest; gastric cancer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Hif-1α Inhibitors Could Successfully Inhibit the Progression of Differentiated Thyroid Cancer in Vitro.Pharmaceuticals (Basel). 2020 Aug 24;13(9):208. doi: 10.3390/ph13090208. Pharmaceuticals (Basel). 2020. PMID: 32847004 Free PMC article.
-
The novel hypoxia-inducible factor-1α inhibitor IDF-11774 regulates cancer metabolism, thereby suppressing tumor growth.Cell Death Dis. 2017 Jun 1;8(6):e2843. doi: 10.1038/cddis.2017.235. Cell Death Dis. 2017. PMID: 28569777 Free PMC article.
-
Helicobacter pylori Induced Phosphatidylinositol-3-OH Kinase/mTOR Activation Increases Hypoxia Inducible Factor-1α to Promote Loss of Cyclin D1 and G0/G1 Cell Cycle Arrest in Human Gastric Cells.Front Cell Infect Microbiol. 2017 Mar 28;7:92. doi: 10.3389/fcimb.2017.00092. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28401064 Free PMC article.
-
Codelivery of HIF-1α siRNA and Dinaciclib by Carboxylated Graphene Oxide-Trimethyl Chitosan-Hyaluronate Nanoparticles Significantly Suppresses Cancer Cell Progression.Pharm Res. 2020 Sep 17;37(10):196. doi: 10.1007/s11095-020-02892-y. Pharm Res. 2020. Retraction in: Pharm Res. 2022 Dec;39(12):3383-3384. doi: 10.1007/s11095-022-03432-6. PMID: 32944844 Retracted.
-
Hypoxic induction of HIF-1alpha and VEGF expression in head and neck squamous cell carcinoma lines is mediated by stress activated protein kinases.Oral Oncol. 2002 Apr;38(3):251-7. doi: 10.1016/s1368-8375(01)00052-5. Oral Oncol. 2002. PMID: 11978547
Cited by
-
Inhibition of Endothelial HIF-1α by IDF-11774 Attenuates Retinal Neovascularization and Vascular Leakage.Invest Ophthalmol Vis Sci. 2025 Jul 1;66(9):47. doi: 10.1167/iovs.66.9.47. Invest Ophthalmol Vis Sci. 2025. PMID: 40668060 Free PMC article.
-
Novel hypoxia-induced HIF-1αactivation in asthma pathogenesis.Respir Res. 2024 Jul 25;25(1):287. doi: 10.1186/s12931-024-02869-0. Respir Res. 2024. PMID: 39061007 Free PMC article.
References
-
- Dulak A.M., Schumacher S.E., van Lieshout J., Imamura Y., Fox C., Shim B., Ramos A.H., Saksena G., Baca S.C., Baselga J., et al. Gastrointestinal adenocarcinomas of the esophagus, stomach, and colon exhibit distinct patterns of genome instability and oncogenesis. Cancer Res. 2012;72:4383–4393. doi: 10.1158/0008-5472.CAN-11-3893. - DOI - PMC - PubMed
-
- Deng N., Goh L.K., Wang H., Das K., Tao J., Tan I.B., Zhang S., Lee M., Wu J., Lim K.H., et al. A comprehensive survey of genomic alterations in gastric cancer reveals systematic patterns of molecular exclusivity and co-occurrence among distinct therapeutic targets. Gut. 2012;61:673–684. doi: 10.1136/gutjnl-2011-301839. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
