

Temperature sensitivity of detrital photosynthesis
- PMID: 38142363
- PMCID: PMC10921823
- DOI: 10.1093/aob/mcad167
Temperature sensitivity of detrital photosynthesis
Erratum in
-
Correction to: Temperature sensitivity of detrital photosynthesis.Ann Bot. 2024 Mar 8;133(1):213-216. doi: 10.1093/aob/mcae013. Ann Bot. 2024. PMID: 38368532 Free PMC article. No abstract available.
Abstract
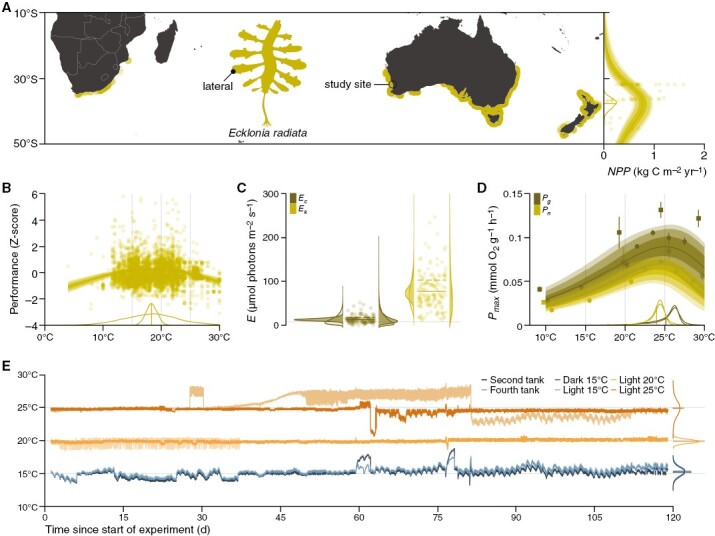
Background and aims: Kelp forests are increasingly considered blue carbon habitats for ocean-based biological carbon dioxide removal, but knowledge gaps remain in our understanding of their carbon cycle. Of particular interest is the remineralization of detritus, which can remain photosynthetically active. Here, we study a widespread, thermotolerant kelp (Ecklonia radiata) to explore detrital photosynthesis as a mechanism underlying temperature and light as two key drivers of remineralization.
Methods: We used meta-analysis to constrain the thermal optimum (Topt) of E. radiata. Temperature and light were subsequently controlled over a 119-day ex situ decomposition experiment. Flow-through experimental tanks were kept in darkness at 15 °C or under a subcompensating maximal irradiance of 8 µmol photons m-2 s-1 at 15, 20 or 25 °C. Photosynthesis of laterals (analogues to leaves) was estimated using closed-chamber oxygen evolution in darkness and under a saturating irradiance of 420 µmol photons m-2 s-1.
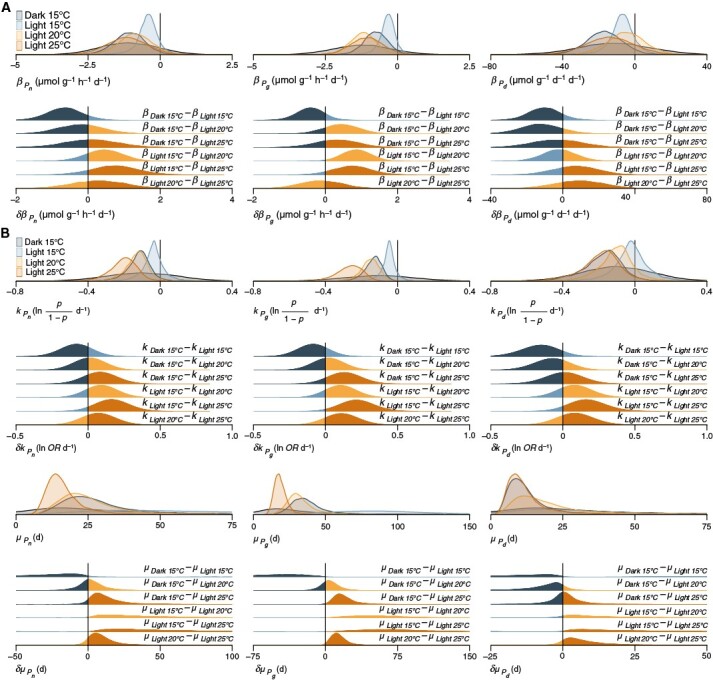
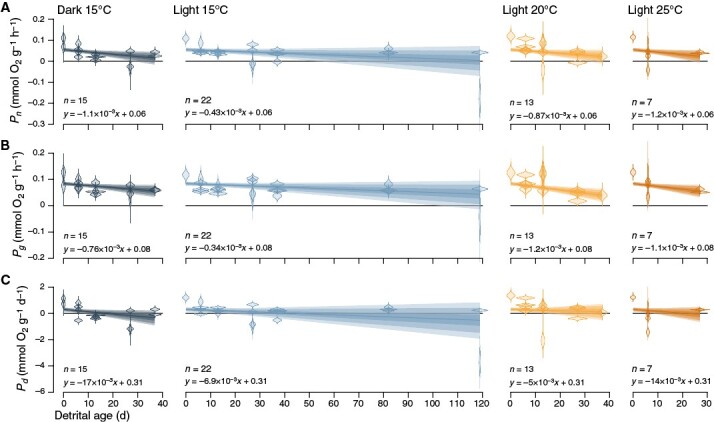
Key results: T opt of E. radiata is 18 °C across performance variables (photosynthesis, growth, abundance, size, mass and fertility), life stages (gametophyte and sporophyte) and populations. Our models predict that a temperature of >15 °C reduces the potential for E. radiata detritus to be photosynthetically viable, hence detrital Topt ≤ 15 °C. Detritus is viable under subcompensating irradiance, where it performs better than in darkness. Comparison of net and gross photosynthesis indicates that elevated temperature primarily decreases detrital photosynthesis, whereas darkness primarily increases detrital respiration compared with optimal experimental conditions, in which detrital photosynthesis can persist for ≥119 days.
Conclusions: T opt of kelp detritus is ≥3 °C colder than that of the intact plant. Given that E. radiata is one of the most temperature-tolerant kelps, this suggests that photosynthesis is generally more thermosensitive in the detrital phase, which partly explains the enhancing effect of temperature on remineralization. In contrast to darkness, even subcompensating irradiance maintains detrital viability, elucidating the accelerating effect of depth and its concomitant light reduction on remineralization to some extent. Detrital photosynthesis is a meaningful mechanism underlying at least two drivers of remineralization, even below the photoenvironment inhabited by the attached alga.
Keywords: Brown algae; CO2; Laminariales; Phaeophyceae; climate change mitigation; detrital dynamics; macroalgae; macroalgal carbon dioxide removal; ocean warming; photoacclimation; photophysiology; temperature tolerance.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Annals of Botany Company.
Figures

References
-
- Andersen GS, Pedersen MF, Nielsen SL.. 2013. Temperature acclimation and heat tolerance of photosynthesis in Norwegian Saccharina latissima (Laminariales, Phaeophyceae). Journal of Phycology 49: 689–700. - PubMed
-
- Arnosti C. 2011. Microbial extracellular enzymes and the marine carbon cycle. Annual Review of Marine Science 3: 401–425. - PubMed
-
- de Bettignies F, Dauby P, Thomas F, et al. 2020. Degradation dynamics and processes associated with the accumulation of Laminaria hyperborea (Phaeophyceae) kelp fragments: an in situ experimental approach. Journal of Phycology 56: 1481–1492. - PubMed
-
- Birch PB, Gabrielson JO, Hamel KS.. 1983. Decomposition of Cladophora. I. Field studies in the Peel-Harvey estuarine system, Western Australia. Botanica Marina 26: 165–172.
-
- Blain CO, Shears NT.. 2019. Seasonal and spatial variation in photosynthetic response of the kelp Ecklonia radiata across a turbidity gradient. Photosynthesis Research 140: 21–38. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
