

Same but different - pseudo-pectin in the charophytic alga Chlorokybus atmophyticus
- PMID: 38148229
- PMCID: PMC10953000
- DOI: 10.1111/ppl.14079
Same but different - pseudo-pectin in the charophytic alga Chlorokybus atmophyticus
Abstract
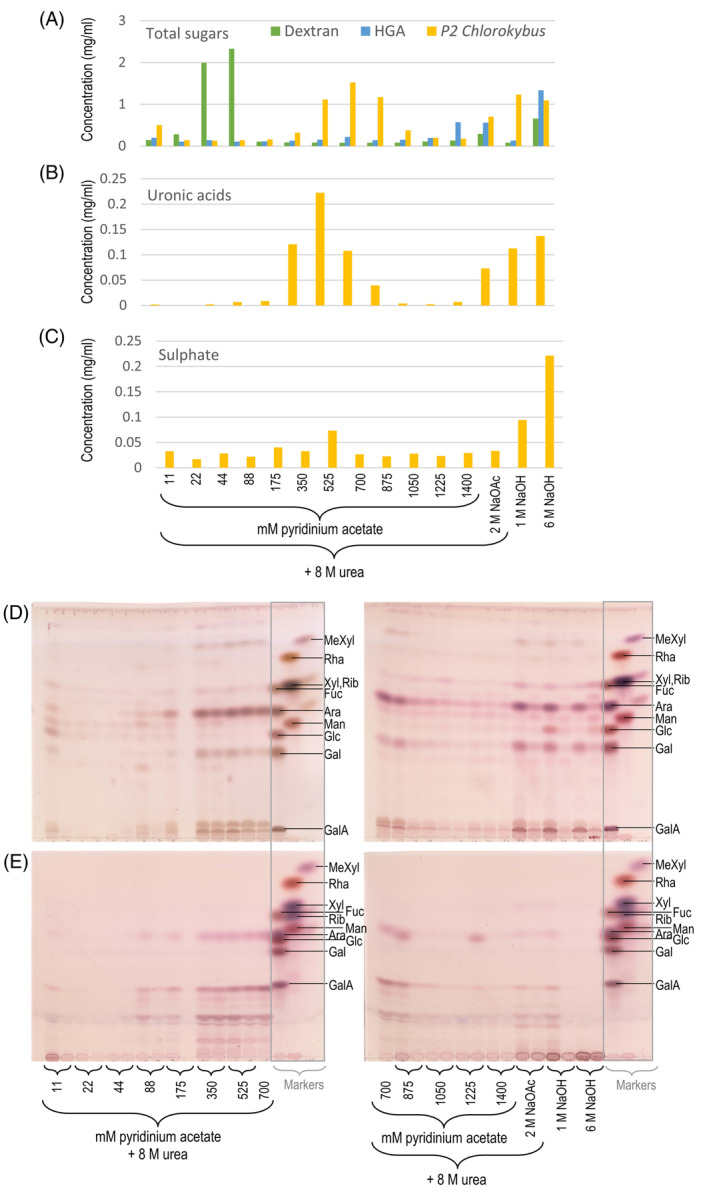
All land-plant cell walls possess hemicelluloses, cellulose and anionic pectin. The walls of their cousins, the charophytic algae, exhibit some similarities to land plants' but also major differences. Charophyte 'pectins' are extractable by conventional land-plant methods, although they differ significantly in composition. Here, we explore 'pectins' of an early-diverging charophyte, Chlorokybus atmophyticus, characterising the anionic polysaccharides that may be comparable to 'pectins' in other streptophytes. Chlorokybus 'pectin' was anionic and upon acid hydrolysis gave GlcA, GalA and sulphate, plus neutral sugars (Ara≈Glc>Gal>Xyl); Rha was undetectable. Most Gal was the l-enantiomer. A relatively acid-resistant disaccharide was characterised as β-d-GlcA-(1→4)-l-Gal. Two Chlorokybus 'pectin' fractions, separable by anion-exchange chromatography, had similar sugar compositions but different sulphate-ester contents. No sugars were released from Chlorokybus 'pectin' by several endo-hydrolases [(1,5)-α-l-arabinanase, (1,4)-β-d-galactanase, (1,4)-β-d-xylanase, endo-polygalacturonase] and exo-hydrolases [α- and β-d-galactosidases, α-(1,6)-d-xylosidase]. 'Driselase', which hydrolyses most land-plant cell wall polysaccharides to mono- and disaccharides, released no sugars except traces of starch-derived Glc. Thus, the Ara, Gal, Xyl and GalA of Chlorokybus 'pectin' were not non-reducing termini with configurations familiar from land-plant polysaccharides (α-l-Araf, α- and β-d-Galp, α- and β-d-Xylp and α-d-GalpA), nor mid-chain residues of α-(1→5)-l-arabinan, β-(1→4)-d-galactan, β-(1→4)-d-xylan or α-(1→4)-d-galacturonan. In conclusion, Chlorokybus possesses anionic 'pectic' polysaccharides, possibly fulfilling pectic roles but differing fundamentally from land-plant pectin. Thus, the evolution of land-plant pectin since the last common ancestor of Chlorokybus and land plants is a long and meandering path involving loss of sulphate, most l-Gal and most d-GlcA; re-configuration of Ara, Xyl and GalA; and gain of Rha.
© 2023 The Authors. Physiologia Plantarum published by John Wiley & Sons Ltd on behalf of Scandinavian Plant Physiology Society.
Figures







References
-
- Albersheim P., Darvill A., Roberts K., Sederoff R., and Staehelin A. (2011) Plant Cell Walls. Garland Science, New York: Taylor and Francis Group.
-
- Anderson C.T. (2016) ‘We be jammin’: an update on pectin biosynthesis, trafficking and dynamics, Journal of Experimental Botany, 67, 495–502. - PubMed
-
- Bischoff H.W. and Bold H.C. (1963) Some soil algae from Enchanted Rock and related algal species. Phycological Studies IV. Univ. Texas Publ., 6318,1‐95.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
