

Cellular and transcriptome signatures unveiled by single-cell RNA-Seq following ex vivo infection of murine splenocytes with Borrelia burgdorferi
- PMID: 38149246
- PMCID: PMC10749944
- DOI: 10.3389/fimmu.2023.1296580
Cellular and transcriptome signatures unveiled by single-cell RNA-Seq following ex vivo infection of murine splenocytes with Borrelia burgdorferi
Abstract
Introduction: Lyme disease, the most common tick-borne infectious disease in the US, is caused by a spirochetal pathogen Borrelia burgdorferi (Bb). Distinct host responses are observed in susceptible and resistant strains of inbred of mice following infection with Bb reflecting a subset of inflammatory responses observed in human Lyme disease. The advent of post-genomic methodologies and genomic data sets enables dissecting the host responses to advance therapeutic options for limiting the pathogen transmission and/or treatment of Lyme disease.
Methods: In this study, we used single-cell RNA-Seq analysis in conjunction with mouse genomics exploiting GFP-expressing Bb to sort GFP+ splenocytes and GFP- bystander cells to uncover novel molecular and cellular signatures that contribute to early stages of immune responses against Bb.
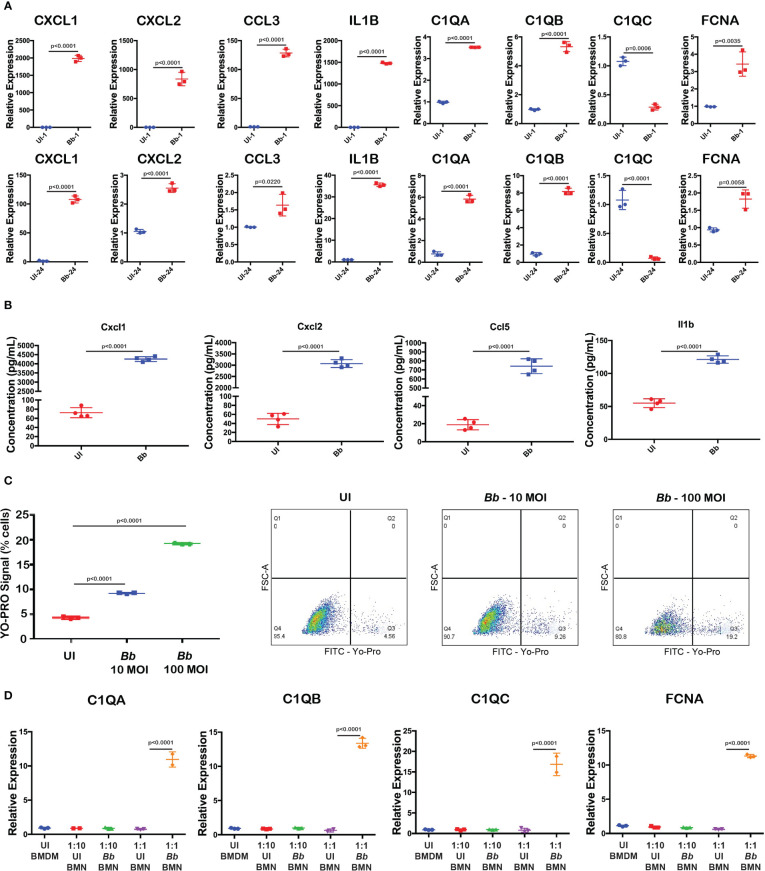
Results: These data decoded the heterogeneity of splenic neutrophils, macrophages, NK cells, B cells, and T cells in C3H/HeN mice in response to Bb infection. Increased mRNA abundance of apoptosis-related genes was observed in neutrophils and macrophages clustered from GFP+ splenocytes. Moreover, complement-mediated phagocytosis-related genes such as C1q and Ficolin were elevated in an inflammatory macrophage subset, suggesting upregulation of these genes during the interaction of macrophages with Bb-infected neutrophils. In addition, the role of DUSP1 in regulating the expression of Casp3 and pro-inflammatory cytokines Cxcl1, Cxcl2, Il1b, and Ccl5 in Bb-infected neutrophils were identified.
Discussion: These findings serve as a growing catalog of cell phenotypes/biomarkers among murine splenocytes that can be exploited for limiting spirochetal burden to limit the transmission of the agent of Lyme disease to humans via reservoir hosts.
Keywords: complement; cytokines; lyme disease; myeloid immune response; neutrophil apoptosis; single-cell RNA-Seq.
Copyright © 2023 Kumaresan, Ingle, Kilgore, Zhang, Hermann and Seshu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Figures









Similar articles
-
Comparative transcriptome analysis of Peromyscus leucopus and C3H mice infected with the Lyme disease pathogen.Front Cell Infect Microbiol. 2023 Apr 11;13:1115350. doi: 10.3389/fcimb.2023.1115350. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37113133 Free PMC article.
-
Ixodes scapularis nymph saliva protein blocks host inflammation and complement-mediated killing of Lyme disease agent, Borrelia burgdorferi.Front Cell Infect Microbiol. 2023 Oct 26;13:1253670. doi: 10.3389/fcimb.2023.1253670. eCollection 2023. Front Cell Infect Microbiol. 2023. PMID: 37965264 Free PMC article.
-
Different patterns of expression and of IL-10 modulation of inflammatory mediators from macrophages of Lyme disease-resistant and -susceptible mice.PLoS One. 2012;7(9):e43860. doi: 10.1371/journal.pone.0043860. Epub 2012 Sep 14. PLoS One. 2012. PMID: 23024745 Free PMC article.
-
The mammalian host response to borrelia infection.Wien Klin Wochenschr. 2006 Nov;118(21-22):653-8. doi: 10.1007/s00508-006-0692-0. Wien Klin Wochenschr. 2006. PMID: 17160603 Review.
-
Investigating disease severity in an animal model of concurrent babesiosis and Lyme disease.Int J Parasitol. 2019 Feb;49(2):145-151. doi: 10.1016/j.ijpara.2018.06.006. Epub 2018 Oct 24. Int J Parasitol. 2019. PMID: 30367867 Free PMC article. Review.
Cited by
-
Changes in the Transcriptome and Long Non-Coding RNAs but Not the Methylome Occur in Human Cells Exposed to Borrelia burgdorferi.Genes (Basel). 2024 Aug 1;15(8):1010. doi: 10.3390/genes15081010. Genes (Basel). 2024. PMID: 39202370 Free PMC article.
-
Multi-Omics Technologies Applied to Improve Tick Research.Microorganisms. 2025 Mar 31;13(4):795. doi: 10.3390/microorganisms13040795. Microorganisms. 2025. PMID: 40284631 Free PMC article. Review.
-
Role of Dual Specificity Phosphatase 1 (DUSP1) in influencing inflammatory pathways in macrophages modulated by Borrelia burgdorferi lipoproteins.bioRxiv [Preprint]. 2024 Nov 21:2024.11.20.624562. doi: 10.1101/2024.11.20.624562. bioRxiv. 2024. PMID: 39605372 Free PMC article. Preprint.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous