

Cyclin D1 extensively reprograms metabolism to support biosynthetic pathways in hepatocytes
- PMID: 38152849
- PMCID: PMC10687208
- DOI: 10.1016/j.jbc.2023.105407
Cyclin D1 extensively reprograms metabolism to support biosynthetic pathways in hepatocytes
Abstract
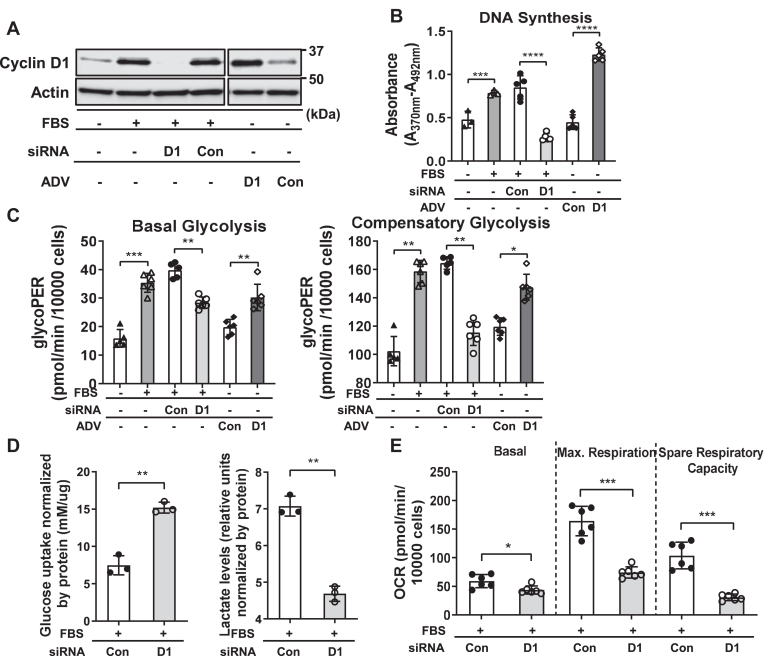
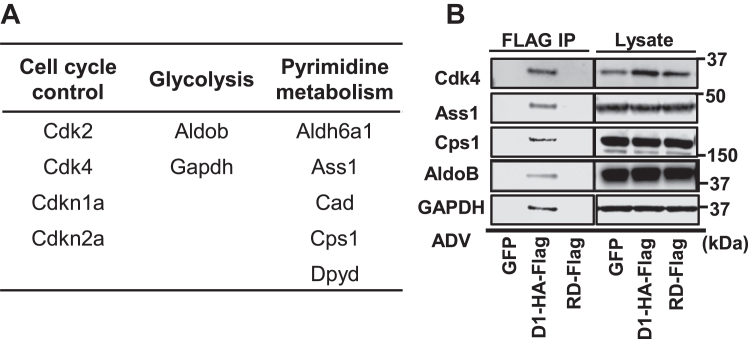
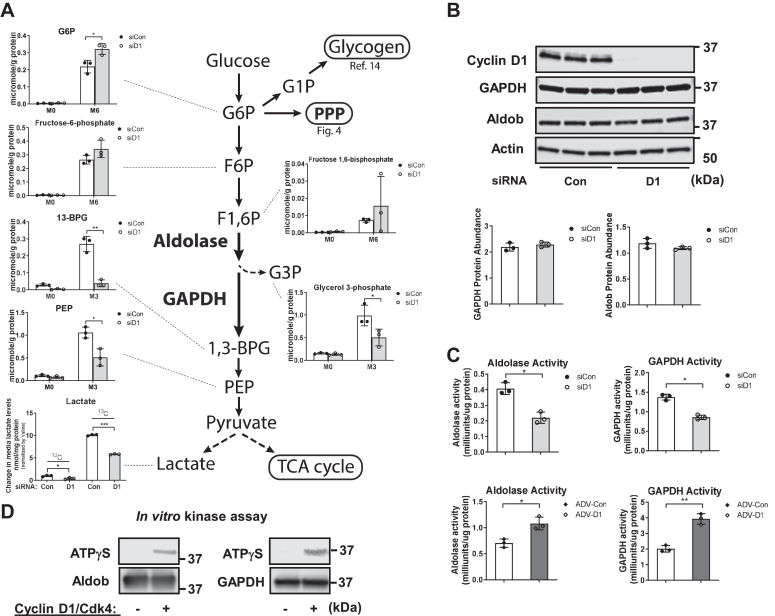
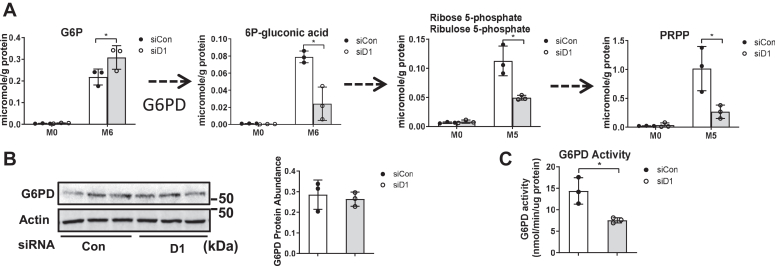
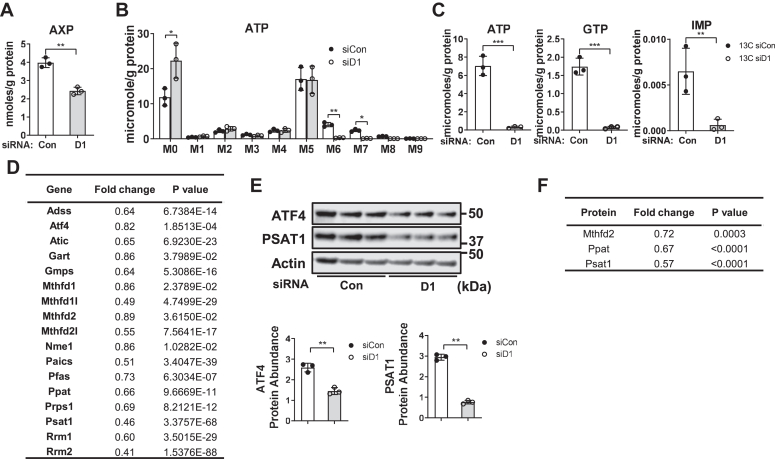
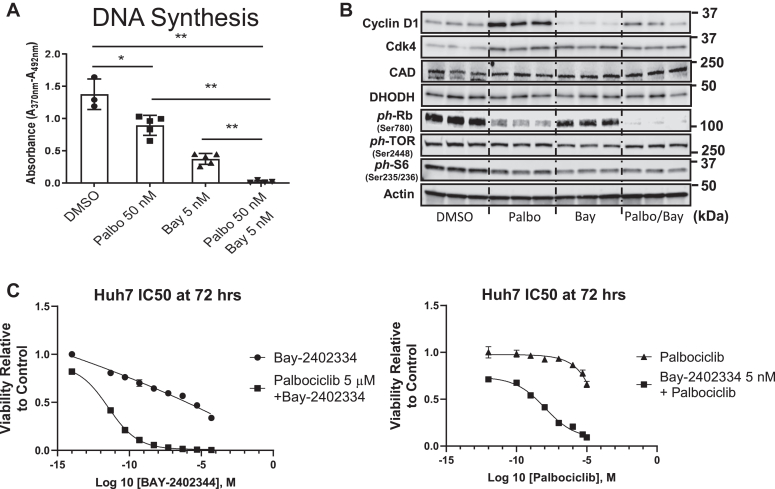
Cell proliferation requires metabolic reprogramming to accommodate biosynthesis of new cell components, and similar alterations occur in cancer cells. However, the mechanisms linking the cell cycle machinery to metabolism are not well defined. Cyclin D1, along with its main partner cyclin-dependent kinase 4 (Cdk4), is a pivotal cell cycle regulator and driver oncogene that is overexpressed in many cancers. Here, we examine hepatocyte proliferation to define novel effects of cyclin D1 on biosynthetic metabolism. Metabolomic studies reveal that cyclin D1 broadly promotes biosynthetic pathways including glycolysis, the pentose phosphate pathway, and the purine and pyrimidine nucleotide synthesis in hepatocytes. Proteomic analyses demonstrate that overexpressed cyclin D1 binds to numerous metabolic enzymes including those involved in glycolysis and pyrimidine synthesis. In the glycolysis pathway, cyclin D1 activates aldolase and GAPDH, and these proteins are phosphorylated by cyclin D1/Cdk4 in vitro. De novo pyrimidine synthesis is particularly dependent on cyclin D1. Cyclin D1/Cdk4 phosphorylates the initial enzyme of this pathway, carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase (CAD), and metabolomic analysis indicates that cyclin D1 depletion markedly reduces the activity of this enzyme. Pharmacologic inhibition of Cdk4 along with the downstream pyrimidine synthesis enzyme dihydroorotate dehydrogenase synergistically inhibits proliferation and survival of hepatocellular carcinoma cells. These studies demonstrate that cyclin D1 promotes a broad network of biosynthetic pathways in hepatocytes, and this model may provide insights into potential metabolic vulnerabilities in cancer cells.
Keywords: BAY 2402234; aldolase; anaerobic glycolysis; cell cycle; cyclin D1; glyceraldehyde-3-phosphate dehydrogenase (GAPDH); liver regeneration; palbociclib; pentose phosphate pathway (PPP); purine; pyrimidine.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures

Similar articles
-
Cyclin D1 inhibits hepatic lipogenesis via repression of carbohydrate response element binding protein and hepatocyte nuclear factor 4α.Cell Cycle. 2012 Jul 15;11(14):2681-90. doi: 10.4161/cc.21019. Epub 2012 Jul 15. Cell Cycle. 2012. PMID: 22751438 Free PMC article.
-
Cyclin D1-Cdk4 controls glucose metabolism independently of cell cycle progression.Nature. 2014 Jun 26;510(7506):547-51. doi: 10.1038/nature13267. Epub 2014 May 25. Nature. 2014. PMID: 24870244 Free PMC article.
-
A negative reciprocal regulatory axis between cyclin D1 and HNF4α modulates cell cycle progression and metabolism in the liver.Proc Natl Acad Sci U S A. 2020 Jul 21;117(29):17177-17186. doi: 10.1073/pnas.2002898117. Epub 2020 Jul 6. Proc Natl Acad Sci U S A. 2020. PMID: 32631996 Free PMC article.
-
Alterations of pRb1-cyclin D1-cdk4/6-p16(INK4A) pathway in endometrial carcinogenesis.Cancer Lett. 2004 Jan 8;203(1):1-12. doi: 10.1016/j.canlet.2003.09.012. Cancer Lett. 2004. PMID: 14670612 Review.
-
Cyclin D1, cancer progression, and opportunities in cancer treatment.J Mol Med (Berl). 2016 Dec;94(12):1313-1326. doi: 10.1007/s00109-016-1475-3. Epub 2016 Oct 2. J Mol Med (Berl). 2016. PMID: 27695879 Free PMC article. Review.
Cited by
-
Autoinducer-2 enhances the defense of Vibrio furnissii against oxidative stress and DNA damage by modulation of c-di-GMP signaling via a two-component system.mBio. 2025 Feb 5;16(2):e0292224. doi: 10.1128/mbio.02922-24. Epub 2025 Jan 16. mBio. 2025. PMID: 39817743 Free PMC article.
-
Patient-derived organotypic tissue cultures as a platform to evaluate metabolic reprogramming in breast cancer patients.J Biol Chem. 2025 May;301(5):108495. doi: 10.1016/j.jbc.2025.108495. Epub 2025 Apr 8. J Biol Chem. 2025. PMID: 40209948 Free PMC article.
-
Hijacking of nucleotide biosynthesis and deamidation-mediated glycolysis by an oncogenic herpesvirus.Nat Commun. 2024 Feb 16;15(1):1442. doi: 10.1038/s41467-024-45852-5. Nat Commun. 2024. PMID: 38365882 Free PMC article.
References
-
- Hanahan D., Weinberg R.A. Hallmarks of cancer: the next generation. Cell. 2011;144:646–674. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
